
13.4: Integration of Somatic Functions
- Page ID
- 22344

By the end of this section, you will be able to:
- Describe the pathways that sensory systems follow into the central nervous system
- Differentiate between the two major ascending pathways in the spinal cord
- Describe the pathway of somatosensory input from the face and compare it to the ascending pathways in the spinal cord
- Explain topographical representations of sensory information
Sensory Nerves
Once any sensory cell transduces a stimulus into a nerve impulse, that impulse has to travel along axons to reach the CNS. In many of the special senses, the axons leaving the sensory receptors have a topographical arrangement, meaning that the location of the sensory receptor relates to the location of the axon in the nerve. For example, in the retina, axons from RGCs in the fovea are located at the center of the optic nerve, where they are surrounded by axons from the more peripheral RGCs.
Spinal Nerves
Generally, spinal nerves contain afferent axons from sensory receptors in the periphery, such as from the skin, mixed with efferent axons travelling to the muscles or other effector organs. As the spinal nerve nears the spinal cord, it splits into dorsal and ventral roots. The dorsal root contains only the axons of sensory neurons, whereas the ventral roots contain only the axons of the motor neurons. Some of the branches will synapse with local neurons in the dorsal root ganglion, posterior (dorsal) horn, or even the anterior (ventral) horn, at the level of the spinal cord where they enter. Other branches will travel a short distance up or down the spine to interact with neurons at other levels of the spinal cord. A branch may also turn into the posterior (dorsal) column of the white matter to connect with the brain. Typically, spinal nerve systems that connect to the brain are contralateral, in that the right side of the body is connected to the left side of the brain and the left side of the body to the right side of the brain.
Cranial Nerves
Cranial nerves convey specific sensory information from the head and neck directly to the brain. For sensations below the neck, the right side of the body is connected to the left side of the brain and the left side of the body to the right side of the brain. Whereas spinal information is contralateral, cranial nerve systems are mostly ipsilateral, meaning that a cranial nerve on the right side of the head is connected to the right side of the brain. Some cranial nerves contain only sensory axons, such as the olfactory, optic, and vestibulocochlear nerves. Other cranial nerves contain both sensory and motor axons, including the trigeminal, facial, glossopharyngeal, and vagus nerves (however, the vagus nerve is not associated with the somatic nervous system). The general senses of somatosensation for the face travel through the trigeminal system.
There are few cranial nerves that are not ipsilateral: optic nerve (CN II), trochlear nerve (CN IV), facial nerve (CN VII), hypoglossal nerves (CN XII). Of these, the optic nerve and facial nerve carry sensory information, while the others carry motor information only.
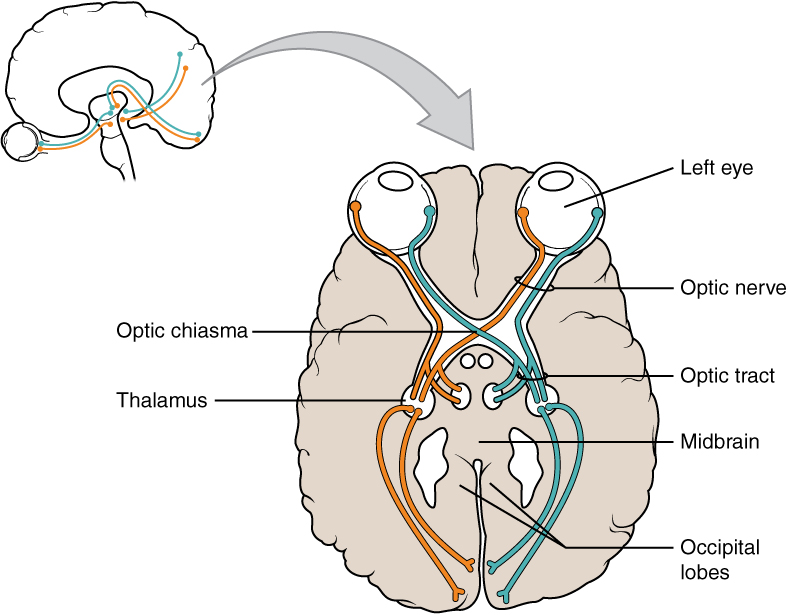
The connections of the optic nerve are more complicated than those of other cranial nerves. Instead of the connections being between each eye and the brain, visual information is segregated between the left and right sides of the visual field. In addition, some of the information from one side of the visual field projects to the opposite side of the brain. Within each eye, the axons projecting from the medial side of the retina cross at the optic chiasm and create new bundles called the optic tract which synapse onto the thalamus and extend to occipital lobes. For example, the axons from the medial retina of the left eye cross over to the right side of the brain at the optic chiasm. However, within each eye, the axons projecting from the lateral side of the retina do not cross. For example, the axons from the lateral retina of the right eye project back to the right side of the brain. Therefore the left field of view of each eye is processed on the right side of the brain, whereas the right field of view of each eye is processed on the left side of the brain (Figure \(\PageIndex{1}\)).
DISORDERS OF THE...
Vision: Bilateral hemianopia
A unique clinical presentation that relates to this anatomic arrangement is the loss of lateral peripheral vision, known as bilateral hemianopia. This is different from “tunnel vision” because the superior and inferior peripheral fields are not lost. Visual field deficits can be disturbing for a patient, but in this case, the cause is not within the visual system itself. A growth of the pituitary gland presses against the optic chiasm and interferes with signal transmission. However, the axons projecting to the same side of the brain are unaffected. Therefore, the patient loses the outermost areas of their field of vision and cannot see objects to their right and left.
Sensory Pathways
Specific regions of the CNS coordinate different somatic processes using sensory inputs and motor outputs of peripheral nerves. A simple case is a reflex caused by a synapse between a dorsal sensory neuron axon and a motor neuron in the ventral horn. More complex arrangements are possible to integrate peripheral sensory information with higher processes. The important regions of the CNS that play a role in somatic processes can be separated into the spinal cord, brainstem, diencephalon, and cerebrum (cortex and subcortical structures).
Spinal Cord and Brainstem
A sensory pathway that carries peripheral sensations to the brain is referred to as an ascending pathway, or ascending tract. Various sensory modalities follow specific pathways through the CNS. Tactile and other somatosensory stimuli activate receptors in the skin, muscles, tendons, and joints throughout the entire body. However, the somatosensory pathways are divided into two separate systems on the basis of the location of the receptor neurons. Somatosensory stimuli from below the neck pass along the sensory pathways of the spinal cord, whereas somatosensory stimuli from the head and neck travel through the cranial nerves—specifically, the trigeminal system.
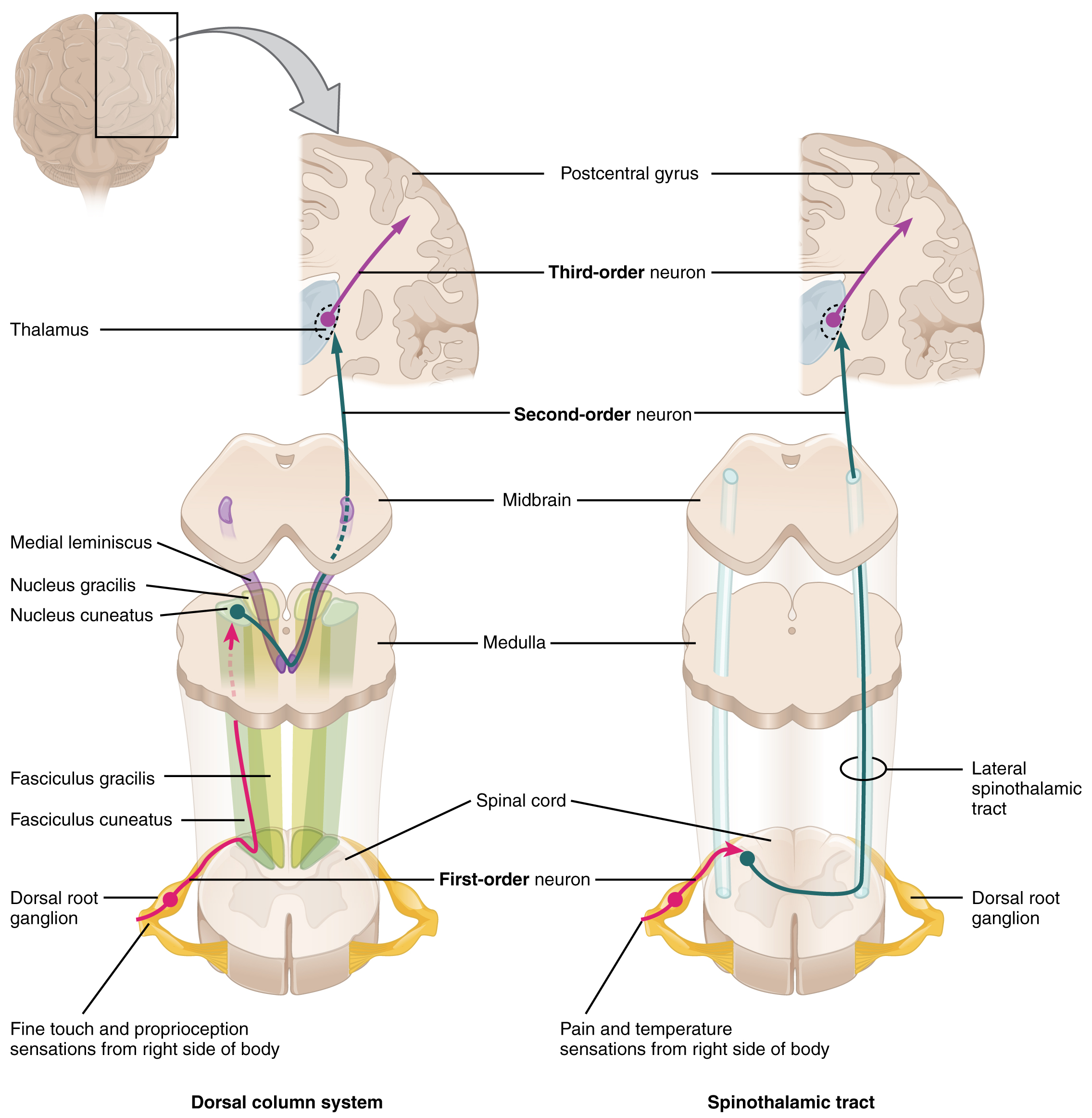
The dorsal column system (sometimes referred to as the dorsal column–medial lemniscus) and the spinothalamic tract are two major pathways that bring sensory information to the brain (Figure \(\PageIndex{2}\)). The sensory pathways in each of these systems are composed of three successive neurons.
The dorsal column system begins with the axon of a dorsal root ganglion neuron entering the dorsal root and joining the dorsal column white matter in the spinal cord. As axons of this pathway enter the dorsal column, they take on a positional arrangement so that axons from lower levels of the body position themselves medially, whereas axons from upper levels of the body position themselves laterally. The dorsal column is separated into two component tracts, the fasciculus gracilis that contains axons from the legs and lower body, and the fasciculus cuneatus that contains axons from the upper body and arms.
The axons in the dorsal column terminate in the nuclei of the medulla, where each synapses with the second neuron in their respective pathway. The nucleus gracilis is the target of fibers in the fasciculus gracilis, whereas the nucleus cuneatus is the target of fibers in the fasciculus cuneatus. The second neuron in the system projects from one of the two nuclei and then decussates, or crosses the midline, in the medulla. These axons then continue to ascend the brainstem as a bundle called the medial lemniscus. These axons terminate in the thalamus, where each synapses with the third neuron in their respective pathway. The third neuron in the system projects its axons to the postcentral gyrus of the cerebral cortex, where somatosensory stimuli are initially processed and the conscious perception of the stimulus occurs.
The spinothalamic tract also begins with neurons in a dorsal root ganglion. These neurons extend their axons to the dorsal horn, where they synapse with the second neuron in their respective pathway. The name “spinothalamic” comes from this second neuron, which has its cell body in the spinal cord gray matter and connects to the thalamus. Axons from these second neurons then decussate within the spinal cord and ascend to the brain and enter the thalamus, where each synapses with the third neuron in its respective pathway. The neurons in the thalamus then project their axons to the spinothalamic tract, which synapses in the postcentral gyrus of the cerebral cortex.
These two systems are similar in that they both begin with dorsal root ganglion cells, as with most general sensory information. The dorsal column system is primarily responsible for touch sensations and proprioception, whereas the spinothalamic tract pathway is primarily responsible for pain and temperature sensations. Another similarity is that the second neurons in both of these pathways project across the midline to the other side of the brain or spinal cord, so that sensation is perceived contralaterally. In the dorsal column system, this decussation takes place in the brainstem; in the spinothalamic pathway, it takes place in the spinal cord at the same spinal cord level at which the information entered. The third neurons in the two pathways are essentially the same. In both, the second neuron synapses in the thalamus, and the thalamic neuron projects to the somatosensory cortex.
The trigeminal pathway carries somatosensory information from the face, head, mouth, and nasal cavity. As with the previously discussed nerve tracts, the sensory pathways of the trigeminal pathway each involve three successive neurons. First, axons from the trigeminal ganglion enter the brainstem at the level of the pons. These axons project to one of three locations. The spinal trigeminal nucleus of the medulla receives information similar to that carried by spinothalamic tract, such as pain and temperature sensations. Other axons go to either the chief sensory nucleus in the pons or the mesencephalic nuclei in the midbrain. These nuclei receive information like that carried by the dorsal column system, such as touch, pressure, vibration, and proprioception. Axons from the second neuron decussate and ascend to the thalamus along the trigeminothalamic tract. In the thalamus, each axon synapses with the third neuron in its respective pathway. Axons from the third neuron then project from the thalamus to the primary somatosensory cortex of the cerebrum.
The sensory pathway for gustation travels along the facial and glossopharyngeal cranial nerves, which synapse with neurons of the solitary nucleus in the brainstem. Axons from the solitary nucleus then project to the ventral posterior nucleus of the thalamus. Finally, axons from the ventral posterior nucleus project to the gustatory cortex of the cerebral cortex, where taste is processed and consciously perceived.
The sensory pathway for audition travels along the vestibulocochlear nerve, which synapses with neurons in the cochlear nuclei of the superior medulla. Within the brainstem, input from either ear is combined to extract location information from the auditory stimuli. Whereas the initial auditory stimuli received at the cochlea strictly represent the frequency—or pitch—of the stimuli, the locations of sounds can be determined by comparing information arriving at both ears. Auditory processing continues on to a nucleus in the midbrain called the inferior colliculus. Axons from the inferior colliculus project to two locations, the thalamus and the superior colliculus. The medial geniculate nucleus of the thalamus receives the auditory information and then projects that information to the auditory cortex in the temporal lobe of the cerebral cortex. The superior colliculus receives input from the visual and somatosensory systems, as well as the ears, to initiate stimulation of the muscles that turn the head and neck toward the auditory stimulus.
Balance is coordinated through the vestibular system, the nerves of which are composed of axons from the vestibular ganglion that carries information from the utricle, saccule, and semicircular canals. The system contributes to controlling head and neck movements in response to vestibular signals. An important function of the vestibular system is coordinating eye and head movements to maintain visual attention. Most of the axons terminate in the vestibular nuclei of the medulla. Some axons project from the vestibular ganglion directly to the cerebellum, with no intervening synapse in the vestibular nuclei. The cerebellum is primarily responsible for initiating movements on the basis of equilibrium information. Neurons in the vestibular nuclei project their axons to targets in the brainstem. One target is the reticular formation, which influences respiratory and cardiovascular functions in relation to body movements. A second target of the axons of neurons in the vestibular nuclei is the spinal cord, which initiates the spinal reflexes involved with posture and balance. To assist the visual system, fibers of the vestibular nuclei project to the oculomotor, trochlear, and abducens nuclei to influence signals sent along the cranial nerves.
The sensory pathway for vision starts with the optic tract. The optic tract has three major targets, two in the diencephalon and one in the midbrain. The connection between the eyes and diencephalon is demonstrated during development, in which the neural tissue of the retina differentiates from that of the diencephalon by the growth of the secondary vesicles. The connections of the retina into the CNS are a holdover from this developmental association. The majority of the connections of the optic tract are to the thalamus—specifically, the lateral geniculate nucleus. Axons from this nucleus then project to the visual cortex of the cerebrum, located in the occipital lobe. Another target of the optic tract is the superior colliculus. In addition, a very small number of retinal ganglion cells (RGCs) axons project from the optic chiasm to the suprachiasmatic nucleus of the hypothalamus. These RGCs are photosensitive, in that they respond to the presence or absence of light. Unlike the photoreceptors, however, these photosensitive RGCs cannot be used to perceive images. By simply responding to the absence or presence of light, these RGCs can send information about day length. The perceived proportion of sunlight to darkness establishes the circadian rhythm of our bodies, allowing certain physiological events to occur at approximately the same time every day.
Diencephalon
The diencephalon is beneath the cerebrum and includes the thalamus and hypothalamus. In the somatic nervous system, the thalamus is an important relay for communication between the cerebrum and the rest of the nervous system. The hypothalamus has both somatic and autonomic functions. In addition, the hypothalamus communicates with the limbic system, which controls emotions and memory functions.
Sensory input to the thalamus comes from most of the special senses and ascending somatosensory tracts. Each sensory system is relayed through a particular nucleus in the thalamus. The thalamus is a required transfer point for most sensory tracts that reach the cerebral cortex, where conscious sensory perception begins. The one exception to this rule is the olfactory system. The olfactory tract axons from the olfactory bulb project directly to the cerebral cortex, along with the limbic system and hypothalamus.
The thalamus is a collection of several nuclei that can be categorized into three anatomical groups. White matter running through the thalamus defines the three major regions of the thalamus, which are an anterior nucleus, a medial nucleus, and a lateral group of nuclei. The anterior nucleus serves as a relay between the hypothalamus and the emotion and memory-producing limbic system. The medial nuclei serve as a relay for information from the limbic system and basal ganglia to the cerebral cortex. This allows memory creation during learning, but also determines alertness. The special and somatic senses connect to the lateral nuclei, where their information is relayed to the appropriate sensory cortex of the cerebrum.
Cerebrum
As described earlier, many of the sensory axons are positioned in the same way as their corresponding receptor cells in the body. This allows identification of the position of a stimulus on the basis of which receptor cells are sending information. The cerebral cortex also maintains this sensory topography in the particular areas of the cortex that correspond to the position of the receptor cells. This mapping is often depicted using a sensory homunculus.
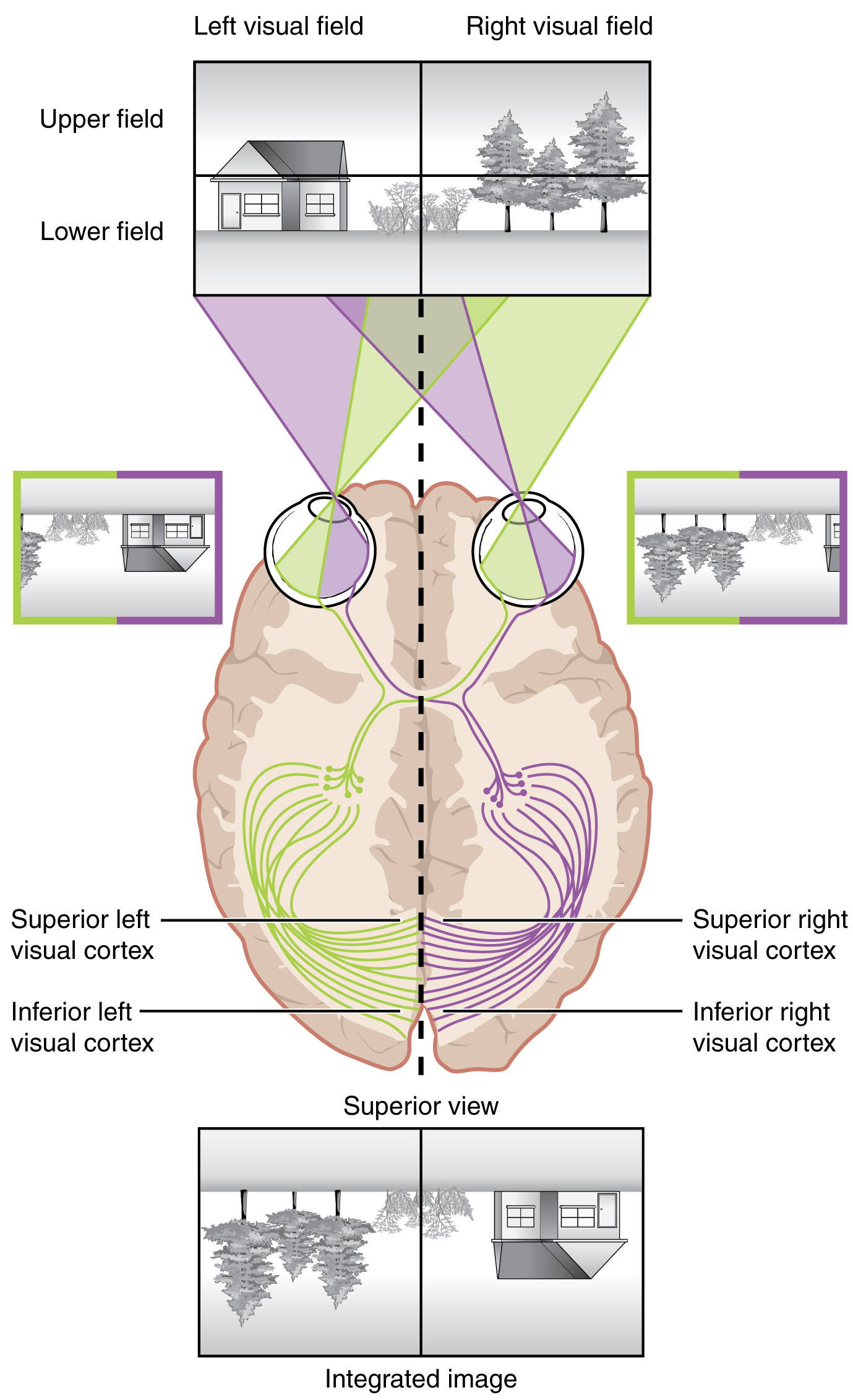
Likewise, the topographic relationship between the retina and the visual cortex is maintained throughout the visual pathway. The visual field is projected onto the two retinae, as described above, with sorting at the optic chiasm. The right peripheral visual field falls on the medial portion of the right retina and the lateral portion of the left retina. The right medial retina then projects across the midline through the optic chiasm. This results in the right visual field being processed in the left visual cortex. Likewise, the left visual field is processed in the right visual cortex. Though the chiasm is helping to sort right and left visual information, superior and inferior visual information is maintained topographically in the visual pathway. Light from the superior visual field falls on the inferior retina, and light from the inferior visual field falls on the superior retina. This topography is maintained such that the superior region of the visual cortex processes the inferior visual field and vice versa. Therefore, the visual field information is inverted and reversed as it enters the visual cortex—up is down, and left is right (Figure \(\PageIndex{3}\)). However, the cortex processes the visual information such that the final conscious perception of the visual field is correct. The topographic relationship is evident in that information from the foveal region of the retina is processed in the center of the primary visual cortex. Information from the peripheral regions of the retina are correspondingly processed toward the edges of the visual cortex. Similar to the exaggerations in the sensory homunculus of the somatosensory cortex, the foveal-processing area of the visual cortex is disproportionately larger than the areas processing peripheral vision.
In an experiment performed in the 1960s, subjects wore prism glasses so that the visual field was inverted before reaching the eye. On the first day of the experiment, subjects would duck when walking up to a table, thinking it was suspended from the ceiling. However, after a few days of acclimation, the subjects behaved as if everything were represented correctly. Therefore, the visual cortex is somewhat flexible in adapting to the information it receives from our eyes.
The cortex has been described as having specific regions that are responsible for processing specific information; there is the visual cortex, somatosensory cortex, gustatory cortex, etc. However, our experience of these senses is not divided. Instead, we experience what can be referred to as a seamless percept. Our perceptions of the various sensory modalities—though distinct in their content—are integrated by the brain so that we experience the world as a continuous whole.
In the cerebral cortex, sensory processing begins at the primary sensory cortex, then proceeds to an association area, and finally, into a multimodal integration area. For example, the visual pathway projects from the retinae through the thalamus to the primary visual cortex in the occipital lobe. This area is primarily in the medial wall within the longitudinal fissure. Here, visual stimuli begin to be recognized as basic shapes. Edges of objects are recognized and built into more complex shapes. Also, inputs from both eyes are compared to extract depth information. Because of the overlapping field of view between the two eyes, the brain can begin to estimate the distance of stimuli based on binocular depth cues.
Concept Review
The nerves that convey sensory information from the periphery to the CNS are either spinal nerves, connected to the spinal cord, or cranial nerves, connected to the brain. Spinal nerves have mixed populations of fibers; some are motor fibers and some are sensory. The sensory fibers connect to the spinal cord through the dorsal root, which is attached to the dorsal root ganglion. Sensory information from the body that is conveyed through spinal nerves will project to the opposite side of the brain to be processed by the cerebral cortex. The cranial nerves can be strictly sensory fibers, such as the olfactory, optic, and vestibulocochlear nerves, or mixed sensory and motor nerves, such as the trigeminal, facial, glossopharyngeal, and vagus nerves. The cranial nerves are connected to the same side of the brain from which the sensory information originates.
Sensory input to the brain enters through pathways that travel through either the spinal cord (for somatosensory input from the body) or the brainstem (for everything else, except the visual and olfactory systems) to reach the diencephalon. In the diencephalon, sensory pathways reach the thalamus. This is necessary for all sensory systems to reach the cerebral cortex, except for the olfactory system that is directly connected to the frontal and temporal lobes.
The two major tracts in the spinal cord, originating from sensory neurons in the dorsal root ganglia, are the dorsal column system and the spinothalamic tract. The major differences between the two are in the type of information that is relayed to the brain and where the tracts decussate. The dorsal column system primarily carries information about touch and proprioception and crosses the midline in the medulla. The spinothalamic tract is primarily responsible for pain and temperature sensation and crosses the midline in the spinal cord at the level at which it enters. The trigeminal nerve adds similar sensation information from the head to these pathways.
The auditory pathway passes through multiple nuclei in the brainstem in which additional information is extracted from the basic frequency stimuli processed by the cochlea. Sound localization is made possible through the activity of these brainstem structures. The vestibular system enters the brainstem and influences activity in the cerebellum, spinal cord, and cerebral cortex.
The visual pathway segregates information from the two eyes so that one half of the visual field projects to the other side of the brain. Within visual cortical areas, the perception of the stimuli and their location is passed along two streams, one ventral and one dorsal. The ventral visual stream connects to structures in the temporal lobe that are important for long-term memory formation. The dorsal visual stream interacts with the somatosensory cortex in the parietal lobe, and together they can influence the activity in the frontal lobe to generate movements of the body in relation to visual information.
Review Questions
Q. Which of these sensory modalities does not pass through the ventral posterior thalamus?
A. gustatory
B. proprioception
C. audition
D. nociception
- Answer
-
C
Q. Which nucleus in the medulla is connected to the inferior colliculus?
A. solitary nucleus
B. vestibular nucleus
C. chief sensory nucleus
D. cochlear nucleus
- Answer
-
D
Q. Visual stimuli in the upper-left visual field will be processed in what region of the primary visual cortex?
A. inferior right
B. inferior left
C. superior right
D. superior left
- Answer
-
A
Q. Which of the following is a direct target of the vestibular ganglion?
A. superior colliculus
B. cerebellum
C. thalamus
D. optic chiasm
- Answer
-
B
Critical Thinking Questions
Q. Following a motorcycle accident, the victim loses the ability to move the right leg but has normal control over the left one, suggesting a hemisection somewhere in the thoracic region of the spinal cord. What sensory deficits would be expected in terms of touch versus pain? Explain your answer.
A. The right leg would feel painful stimuli, but not touch, because the spinothalamic tract decussates at the level of entry, which would be below the injury, whereas the dorsal column system does not decussate until reaching the brainstem, which would be above the injury and thus those fibers would be damaged.
Q. A pituitary tumor can cause perceptual losses in the lateral visual field. The pituitary gland is located directly inferior to the hypothalamus. Why would this happen?
A. As the tumor enlarges, it would press against the optic chiasm, and fibers from the medial retina would be disrupted. These fibers carry information about the lateral visual field because the visual scene is reversed as the light passes through the pupil and lens.
Glossary
- anterior nucleus
- collection of nuclei of the anterior region of the thalamus
- ascending pathway
- fiber structure that relays sensory information from the periphery through the spinal cord and brainstem to other structures of the brain
- association area
- region of cortex connected to a primary sensory cortical area that further processes the information to generate more complex sensory perceptions
- binocular depth cues
- indications of the distance of visual stimuli on the basis of slight differences in the images projected onto either retina
- chief sensory nucleus
- component of the trigeminal nuclei that is found in the pons
- circadian rhythm
- internal perception of the daily cycle of light and dark based on retinal activity related to sunlight
- cochlear nuclei
- nuclei that receive first-order auditory input from the organ of Corti in the cochlea of the inner ear
- contralateral
- word meaning “on the opposite side,” as in axons that cross the midline in a fiber tract
- decussate
- to cross the midline, as in fibers that project from one side of the body to the other
- dorsal column system
- ascending tract of the spinal cord associated with fine touch and proprioceptive sensations
- fasciculus cuneatus
- lateral division of the dorsal column system composed of fibers from sensory neurons in the upper body
- fasciculus gracilis
- medial division of the dorsal column system composed of fibers from sensory neurons in the lower body
- hypothalamus
- major region of the diencephalon that is responsible for coordinating autonomic and endocrine control of homeostasis
- inferior colliculus
- last structure in the auditory brainstem pathway that projects to the thalamus and superior colliculus
- ipsilateral
- word meaning on the same side, as in axons that do not cross the midline in a fiber tract
- lateral geniculate nucleus
- thalamic target of the RGCs that projects to the visual cortex
- lateral group of nuclei
- collection of nuclei of the lateral region of the thalamus
- medial geniculate nucleus
- thalamic target of the auditory brainstem that projects to the auditory cortex
- medial lemniscus
- fiber tract of the dorsal column system that extends from the nuclei gracilis and cuneatus to the thalamus, and decussates
- medial nucleus
- collection of nuclei of the medial region of the thalamus
- mesencephalic nucleus
- component of the trigeminal nuclei that is found in the midbrain
- multimodal integration area
- region of the cerebral cortex in which information from more than one sensory modality is processed to arrive at higher level cortical functions such as memory, learning, or cognition
- nucleus cuneatus
- medullary nucleus at which first-order neurons of the dorsal column system synapse specifically from the upper body and arms
- nucleus gracilis
- medullary nucleus at which first-order neurons of the dorsal column system synapse specifically from the lower body and legs
- optic chiasm
- decussation point in the visual system at which medial retina fibers cross to the other side of the brain
- optic tract
- name for the fiber structure containing axons from the retina posterior to the optic chiasm representing their CNS location
- primary sensory cortex
- region of the cerebral cortex that initially receives sensory input from an ascending pathway from the thalamus and begins the processing that will result in conscious perception of that modality
- solitary nucleus
- medullar nucleus that receives taste information from the facial and glossopharyngeal nerves
- spinal nerve
- one of 31 nerves connected to the spinal cord
- spinal trigeminal nucleus
- component of the trigeminal nuclei that is found in the medulla
- spinothalamic tract
- ascending tract of the spinal cord associated with pain and temperature sensations
- superior colliculus
- structure in the midbrain that combines visual, auditory, and somatosensory input to coordinate spatial and topographic representations of the three sensory systems
- suprachiasmatic nucleus
- hypothalamic target of the retina that helps to establish the circadian rhythm of the body on the basis of the presence or absence of daylight
- thalamus
- major region of the diencephalon that is responsible for relaying information between the cerebrum and the hindbrain, spinal cord, and periphery
- topographical
- relating to positional information
- ventral posterior nucleus
- nucleus in the thalamus that is the target of gustatory sensations and projects to the cerebral cortex
- vestibular nuclei
- targets of the vestibular component of the eighth cranial nerve
Contributors and Attributions
OpenStax Anatomy & Physiology (CC BY 4.0). Access for free at https://openstax.org/books/anatomy-and-physiology