
7.4: Cerebellum and Error-Driven Learning
- Page ID
- 12606

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)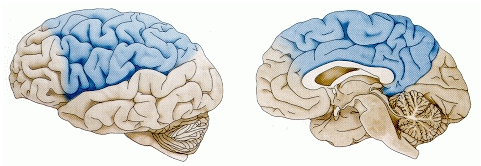
Now that we understand how the basal ganglia can select an action to perform based on reinforcement learning, we turn to the cerebellum, which takes over once the action has been initiated, and uses error-driven learning to shape the performance of the action so that it is accurate and well-coordinated. As shown in Figure 7.10, the cerebellum only receives from cortical areas directly involved in motor production, including the parietal cortex and the motor areas of frontal cortex. Unlike the basal ganglia, it does not receive from prefrontal cortex or temporal cortex, which makes sense according to their respective functions. Prefrontal cortex and temporal cortex are really important for high-level planning and action selection, but not for action execution. However, we do know from neuroimaging experiments that the cerebellum is engaged in many cognitive tasks -- this must reflect its extensive connectivity with the parietal cortex, which is also activated in many cognitive tasks. One idea is that the cerebellum can help shape learning and processing in parietal cortex by virtue of its powerful error-driven learning mechanisms -- this may help to explain how the parietal cortex can learn to do all the complex things it does. However, at this point both the parietal cortex and cerebellum are much better understood from a motor standpoint than a cognitive one.


The cerebellum has a very well-defined anatomy (Figure 7.11, Figure 7.12), with the same basic circuit replicated throughout. Thus, like the basal ganglia, it seems to be performing the same basic function replicated over a wide range of different content domains (e.g., for different motor effectors, and for different areas of parietal and frontal cortex). The basic circuit involves input signals coming from various sources, which are conveyed into the cerebellum via mossy fiber axons. These terminate onto granule cells, of which there are roughly 40 billion in the human brain! Each granule cell receives only 4-5 mossy fiber inputs, and there are roughly 200 million mossy fiber inputs, with each mossy fiber synapsing on roughly 500 granule cells. Thus, there is a great expansion of information coding in the granule cells relative to the input -- we'll revisit this important fact in a moment. To complete the circuit, the granule cells send out parallel fiber axons, that synapse into the very dense dendritic trees of Purkinje cells, which can receive as many as 200,000 inputs from granule cells. There are roughly 15 million Purkinje cells in the human brain, and these cells produce the output signal from the cerebellum. Thus, there is a massive convergence operation from the granule cells onto the Purkinje cells. The Purkinje cells are tonically active, and the granule cells are excitatory onto them, making it a bit puzzling to figure out how the granule cells convey a useful signal to the Purkinje cells. The other cell types in the cerebellum (stellate, basket, and golgi) are inhibitory interneurons that provide inhibitory control over both granule cell and Purkinje cell firing. It is possible that granule cells work in concert with these inhibitory cells to alter the balance of excitation and inhibition in the Purkinje's, but this remains somewhat unclear.
The final piece of the cerebellar puzzle is the climbing fiber input from the inferior olivary nucleus -- there is exactly one such climbing fiber per Purkinje cell, and it has a very powerful effect on the neuron, producing a series of complex spikes. It is thought that climbing fiber inputs convey a training or error signal to the Purkinje's, which then drives synaptic plasticity in its associated granule cell inputs. One prominent idea is that this synaptic plasticity tends to produce LTD (weight decrease) for synaptic inputs where the granule cells are active, which then makes these neurons less likely to fire the Purkinje cell in the future. This would make sense given that the Purkinje cells are inhibitory on the deep cerebellar nuclei neurons, so to produce an output from them, the Purkinje cell needs to be turned off.
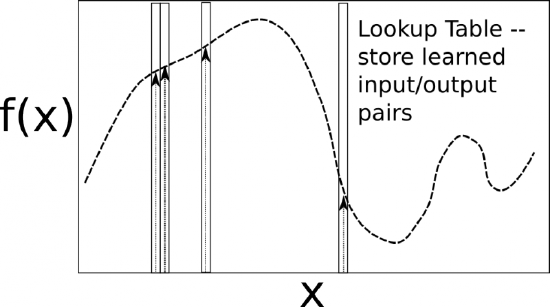
Putting all these pieces together, David Marr and James Albus argued that the cerebellum is a system for error-driven learning, with the error signal coming from the climbing fibers. It is clear that it has the machinery to associate stimulus inputs with motor output commands, under the command of the climbing fiber inputs. One important principle of cerebellar function is the projection of inputs into a very high-dimensional space over the granule cells -- computationally this achieves the separation form of learning, where each combination of inputs activates a unique pattern of granule cell neurons. This unique pattern can then be associated with a different output signal from the cerebellum, producing something approximating a lookup table of input/output values Figure 7.13. A lookup table provides a very robust solution to learning even very complex, arbitrary functions -- it will always be able to encode any kind of function. The drawback is that it does not generalize to novel input patterns very well. However, it may be better overall in motor control to avoid improper generalization, rather than eek out a bit more efficiency from some form of generalization. This high-dimensional expansion is also used successfully by the support vector machine (SVM), one of the most successful machine learning algorithms.
{kind=link}
Exploration of Cerebellum
- Cereb (cereb.proj) -- Cerebellum role in motor learning, learning from errors.