
8.3: Special Senses
- Page ID
- 9585

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Explain how olfactory, gustatory, equilibrium, auditory, and visual sensations are interpreted by animals with reference to signal processing and/or specific ion movements.
Sensory information processing in animals
In more advanced animals, the senses are constantly at work, making the animal aware of stimuli—such as light, or sound, or the presence of a chemical substance in the external environment—and monitoring information about the organism’s internal environment. All bilaterally symmetric animals have a sensory system, and the development of any species’ sensory system has been driven by natural selection; thus, sensory systems differ among species according to the demands of their environments. The shark, unlike most fish predators, is electrosensitive—that is, sensitive to electrical fields produced by other animals in its environment. While it is helpful to this underwater predator, electrosensitivity is a sense not found in most land animals.
Senses provide information about the body and its environment. Humans have five special senses: olfaction (smell), gustation (taste), equilibrium (balance and body position), vision, and hearing. Additionally, we possess general senses, also called somatosensation, which respond to stimuli like temperature, pain, pressure, and vibration. Vestibular sensation, which is an organism’s sense of spatial orientation and balance, proprioception (position of bones, joints, and muscles), and the sense of limb position that is used to track kinesthesia (limb movement) are part of somatosensation. Although the sensory systems associated with these senses are very different, all share a common function: to convert a stimulus (such as light, or sound, or the position of the body) into an electrical signal in the nervous system. This process is called sensory transduction.
There are two broad types of cellular systems that perform sensory transduction. In one, a neuron works with a sensory receptor, a cell, or cell process that is specialized to engage with and detect a specific stimulus. Stimulation of the sensory receptor activates the associated afferent neuron, which carries information about the stimulus to the central nervous system. In the second type of sensory transduction, a sensory nerve ending responds to a stimulus in the internal or external environment: this neuron constitutes the sensory receptor. Free nerve endings can be stimulated by several different stimuli, thus showing little receptor specificity. For example, pain receptors in your gums and teeth may be stimulated by temperature changes, chemical stimulation, or pressure.
Reception
The first step in sensation is reception, which is the activation of sensory receptors by stimuli such as mechanical stimuli (being bent or squished, for example), chemicals, or temperature. The receptor can then respond to the stimuli. The region in space in which a given sensory receptor can respond to a stimulus, be it far away or in contact with the body, is that receptor’s receptive field. Think for a moment about the differences in receptive fields for the different senses. For the sense of touch, a stimulus must come into contact with body. For the sense of hearing, a stimulus can be a moderate distance away (some baleen whale sounds can propagate for many kilometers). For vision, a stimulus can be very far away; for example, the visual system perceives light from stars at enormous distances.
Transduction
The most fundamental function of a sensory system is the translation of a sensory signal to an electrical signal in the nervous system. This takes place at the sensory receptor, and the change in electrical potential that is produced is called the receptor potential. How is sensory input, such as pressure on the skin, changed to a receptor potential? In this example, a type of receptor called a mechanoreceptor (as shown in Figure 8.14) possesses specialized membranes that respond to pressure. Disturbance of these dendrites by compressing them or bending them opens gated ion channels in the plasma membrane of the sensory neuron, changing its electrical potential. Recall that in the nervous system, a positive change of a neuron’s electrical potential (also called the membrane potential), depolarizes the neuron. Receptor potentials are graded potentials: the magnitude of these graded (receptor) potentials varies with the strength of the stimulus. If the magnitude of depolarization is sufficient (that is, if membrane potential reaches a threshold), the neuron will fire an action potential. In most cases, the correct stimulus impinging on a sensory receptor will drive membrane potential in a positive direction, although for some receptors, such as those in the visual system, this is not always the case.
Sensory receptors for different senses are very different from each other, and they are specialized according to the type of stimulus they sense: they have receptor specificity. For example, touch receptors, light receptors, and sound receptors are each activated by different stimuli. Touch receptors are not sensitive to light or sound; they are sensitive only to touch or pressure. However, stimuli may be combined at higher levels in the brain, as happens with olfaction, contributing to our sense of taste.
Encoding and transmission of sensory information
Four aspects of sensory information are encoded by sensory systems: the type of stimulus, the location of the stimulus in the receptive field, the duration of the stimulus, and the relative intensity of the stimulus. Thus, action potentials transmitted over a sensory receptor’s afferent axons encode one type of stimulus, and this segregation of the senses is preserved in other sensory circuits. For example, auditory receptors transmit signals over their own dedicated system, and electrical activity in the axons of the auditory receptors will be interpreted by the brain as an auditory stimulus—a sound.
The intensity of a stimulus is often encoded in the rate of action potentials produced by the sensory receptor. Thus, an intense stimulus will produce a more rapid train of action potentials, and reducing the stimulus will likewise slow the rate of production of action potentials. A second way in which intensity is encoded is by the number of receptors activated. An intense stimulus might initiate action potentials in a large number of adjacent receptors, while a less intense stimulus might stimulate fewer receptors. Integration of sensory information begins as soon as the information is received in the CNS, and the brain will further process incoming signals.
Perception
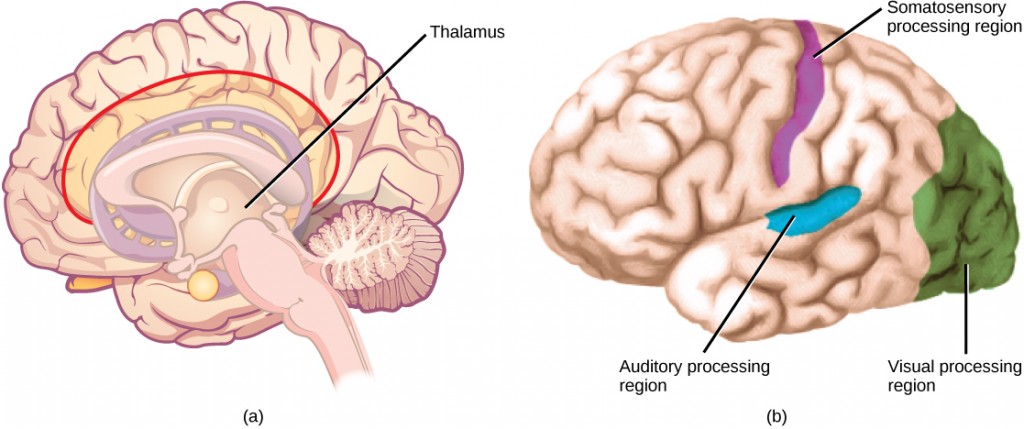
Perception is an individual’s interpretation of a sensation. Although perception relies on the activation of sensory receptors, perception happens not at the level of the sensory receptor, but at higher levels in the nervous system, in the brain. The brain distinguishes sensory stimuli through a sensory pathway: action potentials from sensory receptors travel along neurons that are dedicated to a particular stimulus. These neurons are dedicated to that particular stimulus and synapse with particular neurons in the brain or spinal cord.
All sensory signals, except those from the olfactory system, are transmitted though the central nervous system and are routed to the thalamus and to the appropriate region of the cortex. Recall that the thalamus is a structure in the forebrain that serves as a clearinghouse and relay station for sensory (as well as motor) signals. When the sensory signal exits the thalamus, it is conducted to the specific area of the cortex (Figure 8.15) dedicated to processing that particular sense.
How are neural signals interpreted? Interpretation of sensory signals between individuals of the same species is largely similar, owing to the inherited similarity of their nervous systems; however, there are some individual differences. A good example of this is individual tolerances to a painful stimulus, such as dental pain, which certainly differ.
Where does perception occur?
a. spinal cord
b. cerebral cortex
c. receptors
d. thalamus
If a person’s cold receptors no longer convert cold stimuli into sensory signals, that person has a problem with the process of ________.
a. reception
b. transmission
c. perception
d. transduction
After somatosensory transduction, the sensory signal travels through the brain as a(n) _____ signal.
a. electrical
b. pressure
c. optical
d. thermal
Somatosensation
Somatosensation is a mixed sensory category and includes all sensation received from the skin and mucous membranes, as well from as the limbs and joints. Somatosensation is also known as tactile sense, or more familiarly, as the sense of touch. Somatosensation occurs all over the exterior of the body and at some interior locations as well. A variety of receptor types—embedded in the skin, mucous membranes, muscles, joints, internal organs, and cardiovascular system—play a role.
Somatosensory receptors
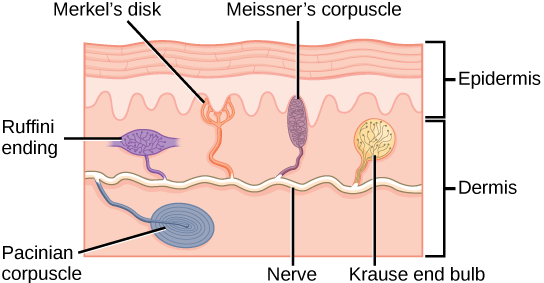
Sensory receptors are classified into five categories: mechanoreceptors, thermoreceptors, proprioceptors, pain receptors, and chemoreceptors. These categories are based on the nature of stimuli each receptor class transduces. What is commonly referred to as “touch” involves more than one kind of stimulus and more than one kind of receptor. Mechanoreceptors in the skin are described as encapsulated (that is, surrounded by a capsule) or unencapsulated (a group that includes free nerve endings). A free nerve ending, as its name implies, is an unencapsulated dendrite of a sensory neuron. Free nerve endings are the most common nerve endings in skin, and they extend into the middle of the epidermis. Free nerve endings are sensitive to painful stimuli, to hot and cold, and to light touch. They are slow to adjust to a stimulus and so are less sensitive to abrupt changes in stimulation.
There are three classes of mechanoreceptors: tactile, proprioceptors, and baroreceptors. Mechanoreceptors sense stimuli due to physical deformation of their plasma membranes. They contain mechanically gated ion channels whose gates open or close in response to pressure, touch, stretching, and sound.” There are four primary tactile mechanoreceptors in human skin: Merkel’s disks, Meissner’s corpuscles, Ruffini endings, and Pacinian corpuscle; two are located toward the surface of the skin and two are located deeper. A fifth type of mechanoreceptor, Krause end bulbs, are found only in specialized regions. Merkel’s disks (shown in Figure 8.16) are found in the upper layers of skin near the base of the epidermis, both in skin that has hair and on glabrous skin, that is, the hairless skin found on the palms and fingers, the soles of the feet, and the lips of humans and other primates. Merkel’s disks are densely distributed in the fingertips and lips. They are slow-adapting, unencapsulated nerve endings, and they respond to light touch. Light touch, also known as discriminative touch, is a light pressure that allows the location of a stimulus to be pinpointed. The receptive fields of Merkel’s disks are small with well-defined borders. That makes them finely sensitive to edges and they come into use in tasks such as typing on a keyboard.
Which of the following statements about mechanoreceptors is false?
a. Pacini corpuscles are found in both glabrous and hairy skin.
b. Merkel’s disks are abundant on the fingertips and lips.
c. Ruffini endings are encapsulated mechanoreceptors.
d. Meissner’s corpuscles extend into the lower dermis.
In proprioception, proprioceptive and kinesthetic signals travel through myelinated afferent neurons running from the spinal cord to the medulla. Neurons are not physically connected, but communicate via neurotransmitters secreted into synapses or “gaps” between communicating neurons. Once in the medulla, the neurons continue carrying the signals to the thalamus.
Muscle spindles are stretch receptors that detect the amount of stretch, or lengthening of muscles. Related to these are Golgi tendon organs, which are tension receptors that detect the force of muscle contraction. Proprioceptive and kinesthetic signals come from limbs. Unconscious proprioceptive signals run from the spinal cord to the cerebellum, the brain region that coordinates muscle contraction, rather than to the thalamus, like most other sensory information.
Barorecptors detect pressure changes in an organ. They are found in the walls of the carotid artery and the aorta where they monitor blood pressure, and in the lungs where they detect the degree of lung expansion. Stretch receptors are found at various sites in the digestive and urinary systems.
In addition to these two types of deeper receptors, there are also rapidly adapting hair receptors, which are found on nerve endings that wrap around the base of hair follicles. There are a few types of hair receptors that detect slow and rapid hair movement, and they differ in their sensitivity to movement. Some hair receptors also detect skin deflection, and certain rapidly adapting hair receptors allow detection of stimuli that have not yet touched the skin.
Integration of signals from mechanoreceptors
The configuration of the different types of receptors working in concert in human skin results in a very refined sense of touch. The nociceptive receptors—those that detect pain—are located near the surface. Small, finely calibrated mechanoreceptors—Merkel’s disks and Meissner’s corpuscles—are located in the upper layers and can precisely localize even gentle touch. The large mechanoreceptors—Pacinian corpuscles and Ruffini endings—are located in the lower layers and respond to deeper touch. (Consider that the deep pressure that reaches those deeper receptors would not need to be finely localized.) Both the upper and lower layers of the skin hold rapidly and slowly adapting receptors. Both primary somatosensory cortex and secondary cortical areas are responsible for processing the complex picture of stimuli transmitted from the interplay of mechanoreceptors.
Density of mechanoreceptors
The distribution of touch receptors in human skin is not consistent over the body. In humans, touch receptors are less dense in skin covered with any type of hair, such as the arms, legs, torso, and face. Touch receptors are denser in glabrous skin (the type found on human fingertips and lips, for example), which is typically more sensitive and is thicker than hairy skin (4 to 5 mm versus 2 to 3 mm).
How is receptor density estimated in a human subject? The relative density of pressure receptors in different locations on the body can be demonstrated experimentally using a two-point discrimination test. In this demonstration, two sharp points, such as two thumbtacks, are brought into contact with the subject’s skin (though not hard enough to cause pain or break the skin). The subject reports if he or she feels one point or two points. If the two points are felt as one point, it can be inferred that the two points are both in the receptive field of a single sensory receptor. If two points are felt as two separate points, each is in the receptive field of two separate sensory receptors. The points could then be moved closer and re-tested until the subject reports feeling only one point, and the size of the receptive field of a single receptor could be estimated from that distance.
Thermoreception
In addition to Krause end bulbs that detect cold and Ruffini endings that detect warmth, there are different types of cold receptors on some free nerve endings: thermoreceptors, located in the dermis, skeletal muscles, liver, and hypothalamus, that are activated by different temperatures. Their pathways into the brain run from the spinal cord through the thalamus to the primary somatosensory cortex. Warmth and cold information from the face travels through one of the cranial nerves to the brain. You know from experience that a tolerably cold or hot stimulus can quickly progress to a much more intense stimulus that is no longer tolerable. Any stimulus that is too intense can be perceived as pain because temperature sensations are conducted along the same pathways that carry pain sensations
Pain
Pain is the name given to nociception, which is the neural processing of injurious stimuli in response to tissue damage. Pain is caused by true sources of injury, such as contact with a heat source that causes a thermal burn or contact with a corrosive chemical. But pain also can be caused by harmless stimuli that mimic the action of damaging stimuli, such as contact with capsaicins, the compounds that cause peppers to taste hot and which are used in self-defense pepper sprays and certain topical medications. Peppers taste “hot” because the protein receptors that bind capsaicin open the same calcium channels that are activated by warm receptors.
Nociception starts at the sensory receptors, but pain, inasmuch as it is the perception of nociception, does not start until it is communicated to the brain. There are several nociceptive pathways to and through the brain. Most axons carrying nociceptive information into the brain from the spinal cord project to the thalamus (as do other sensory neurons) and the neural signal undergoes final processing in the primary somatosensory cortex. Interestingly, one nociceptive pathway projects not to the thalamus but directly to the hypothalamus in the forebrain, which modulates the cardiovascular and neuroendocrine functions of the autonomic nervous system. Recall that threatening—or painful—stimuli stimulate the sympathetic branch of the visceral sensory system, readying a fight-or-flight response.
- View this video that animates the five phases of nociceptive pain.
Taste and smell
Taste, also called gustation, and smell, also called olfaction, are the most interconnected senses in that both involve molecules of the stimulus entering the body and bonding to receptors. Smell lets an animal sense the presence of food or other animals—whether potential mates, predators, or prey—or other chemicals in the environment that can impact their survival. Similarly, the sense of taste allows animals to discriminate between types of foods. While the value of a sense of smell is obvious, what is the value of a sense of taste? Different tasting foods have different attributes, both helpful and harmful. For example, sweet-tasting substances tend to be highly caloric, which could be necessary for survival in lean times. Bitterness is associated with toxicity, and sourness is associated with spoiled food. Salty foods are valuable in maintaining homeostasis by helping the body retain water and by providing ions necessary for cells to function.
Tastes and odors
Both taste and odor stimuli are molecules taken in from the environment. The primary tastes detected by humans are sweet, sour, bitter, salty and umami. The first four tastes need little explanation. The identification of umami as a fundamental taste occurred fairly recently—it was identified in 1908 by Japanese scientist Kikunae Ikeda while he worked with seaweed broth, but it was not widely accepted as a taste that could be physiologically distinguished until many years later. The taste of umami, also known as savoriness, is attributable to the taste of the amino acid L-glutamate. In fact, monosodium glutamate, or MSG, is often used in cooking to enhance the savory taste of certain foods. What is the adaptive value of being able to distinguish umami? Savory substances tend to be high in protein.
All odors that we perceive are molecules in the air we breathe. If a substance does not release molecules into the air from its surface, it has no smell. And if a human or other animal does not have a receptor that recognizes a specific molecule, then that molecule has no smell. Humans have about 350 olfactory receptor subtypes that work in various combinations to allow us to sense about 10,000 different odors. Compare that to mice, for example, which have about 1,300 olfactory receptor types, and therefore probably sense more odors. Both odors and tastes involve molecules that stimulate specific chemoreceptors. Although humans commonly distinguish taste as one sense and smell as another, they work together to create the perception of flavor. A person’s perception of flavor is reduced if he or she has congested nasal passages.
Reception and transduction
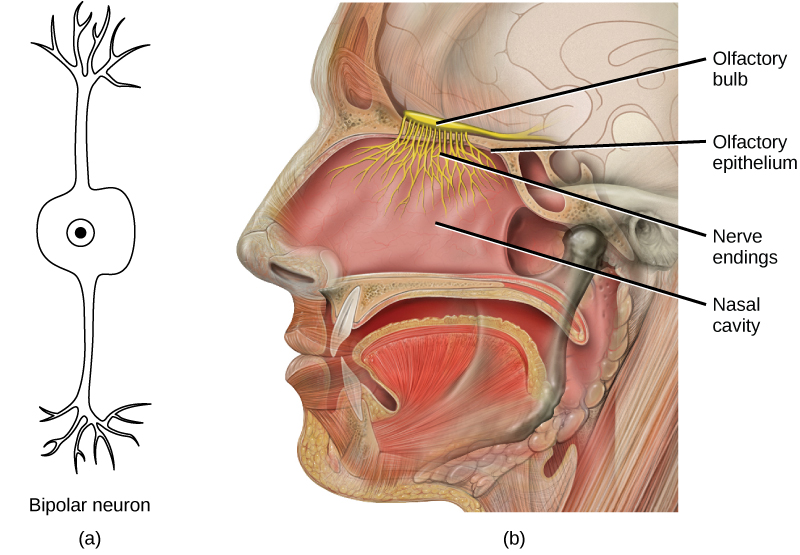
Odorants (odor molecules) enter the nose and dissolve in the olfactory epithelium, the mucosa at the back of the nasal cavity (as illustrated in Figure 8.17). The olfactory epithelium is a collection of specialized olfactory receptors in the back of the nasal cavity that spans an area about 5 cm2 in humans. Recall that sensory cells are neurons. An olfactory receptor, which is a dendrite of a specialized neuron, responds when it binds certain molecules inhaled from the environment by sending impulses directly to the olfactory bulb of the brain. Humans have about 12 million olfactory receptors, distributed among hundreds of different receptor types that respond to different odors. Twelve million seems like a large number of receptors, but compare that to other animals: rabbits have about 100 million, most dogs have about 1 billion, and bloodhounds—dogs selectively bred for their sense of smell—have about 4 billion. The overall size of the olfactory epithelium also differs between species, with that of bloodhounds, for example, being many times larger than that of humans.
Olfactory neurons are bipolar neurons (neurons with two processes from the cell body). Each neuron has a single dendrite buried in the olfactory epithelium, and extending from this dendrite are 5 to 20 receptor-laden, hair-like cilia that trap odorant molecules. The sensory receptors on the cilia are proteins, and it is the variations in their amino acid chains that make the receptors sensitive to different odorants. Each olfactory sensory neuron has only one type of receptor on its cilia, and the receptors are specialized to detect specific odorants, so the bipolar neurons themselves are specialized. When an odorant binds with a receptor that recognizes it, the sensory neuron associated with the receptor is stimulated. Olfactory stimulation is the only sensory information that directly reaches the cerebral cortex, whereas other sensations are relayed through the thalamus.
Pheromones
A pheromone is a chemical released by an animal that affects the behavior or physiology of animals of the same species. Pheromonal signals can have profound effects on animals that inhale them, but pheromones apparently are not consciously perceived in the same way as other odors. There are several different types of pheromones, which are released in urine or as glandular secretions. Certain pheromones are attractants to potential mates, others are repellants to potential competitors of the same sex, and still others play roles in mother-infant attachment. Some pheromones can also influence the timing of puberty, modify reproductive cycles, and even prevent embryonic implantation. While the roles of pheromones in many nonhuman species are important, pheromones have become less important in human behavior over evolutionary time compared to their importance to organisms with more limited behavioral repertoires.
The vomeronasal organ (VNO, or Jacobson’s organ) is a tubular, fluid-filled, olfactory organ present in many vertebrate animals that sits adjacent to the nasal cavity. It is very sensitive to pheromones and is connected to the nasal cavity by a duct. When molecules dissolve in the mucosa of the nasal cavity, they then enter the VNO where the pheromone molecules among them bind with specialized pheromone receptors. Upon exposure to pheromones from their own species or others, many animals, including cats, may display the flehmen response (shown in Figure 8.18), a curling of the upper lip that helps pheromone molecules enter the VNO.
Pheromonal signals are sent, not to the main olfactory bulb, but to a different neural structure that projects directly to the amygdala (recall that the amygdala is a brain center important in emotional reactions, such as fear). The pheromonal signal then continues to areas of the hypothalamus that are key to reproductive physiology and behavior. While some scientists assert that the VNO is apparently functionally vestigial in humans, eve though there is a similar structure located near human nasal cavities, others are researching it as a possible functional system that may, for example, contribute to synchronization of menstrual cycles in women living in close proximity.

Taste
Detecting a taste (gustation) is fairly similar to detecting an odor (olfaction), given that both taste and smell rely on chemical receptors being stimulated by certain molecules. The primary organ of taste is the taste bud. A taste bud is a cluster of gustatory receptors (taste cells) that are located within the bumps on the tongue called papillae (singular: papilla) (illustrated in Figure 8.19). There are several structurally distinct papillae. Filiform papillae, which are located across the tongue, are tactile, providing friction that helps the tongue move substances, and contain no taste cells. In contrast, fungiform papillae, which are located mainly on the anterior two-thirds of the tongue, each contain one to eight taste buds and also have receptors for pressure and temperature. The large circumvallate papillae contain up to 100 taste buds and form a V near the posterior margin of the tongue.
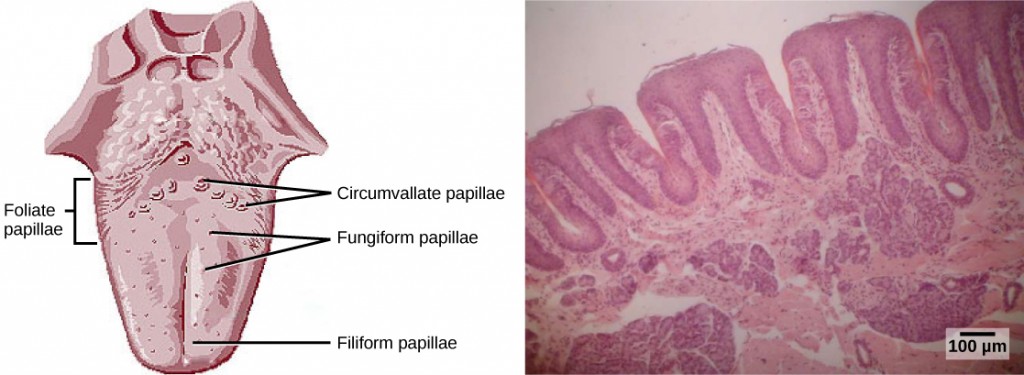
In addition to those two types of chemically and mechanically sensitive papillae are foliate papillae—leaf-like papillae located in parallel folds along the edges and toward the back of the tongue, as seen in the Figure 8.18b micrograph. Foliate papillae contain about 1,300 taste buds within their folds. Finally, there are circumvallate papillae, which are wall-like papillae in the shape of an inverted “V” at the back of the tongue. Each of these papillae is surrounded by a groove and contains about 250 taste buds.
Each taste bud’s taste cells are replaced every 10 to 14 days. These are elongated cells with hair-like processes called microvilli at the tips that extend into the taste bud pore (illustrate in Figure 8.20). Food molecules (tastants) are dissolved in saliva, and they bind with and stimulate the receptors on the microvilli. The receptors for tastants are located across the outer portion and front of the tongue, outside of the middle area where the filiform papillae are most prominent.
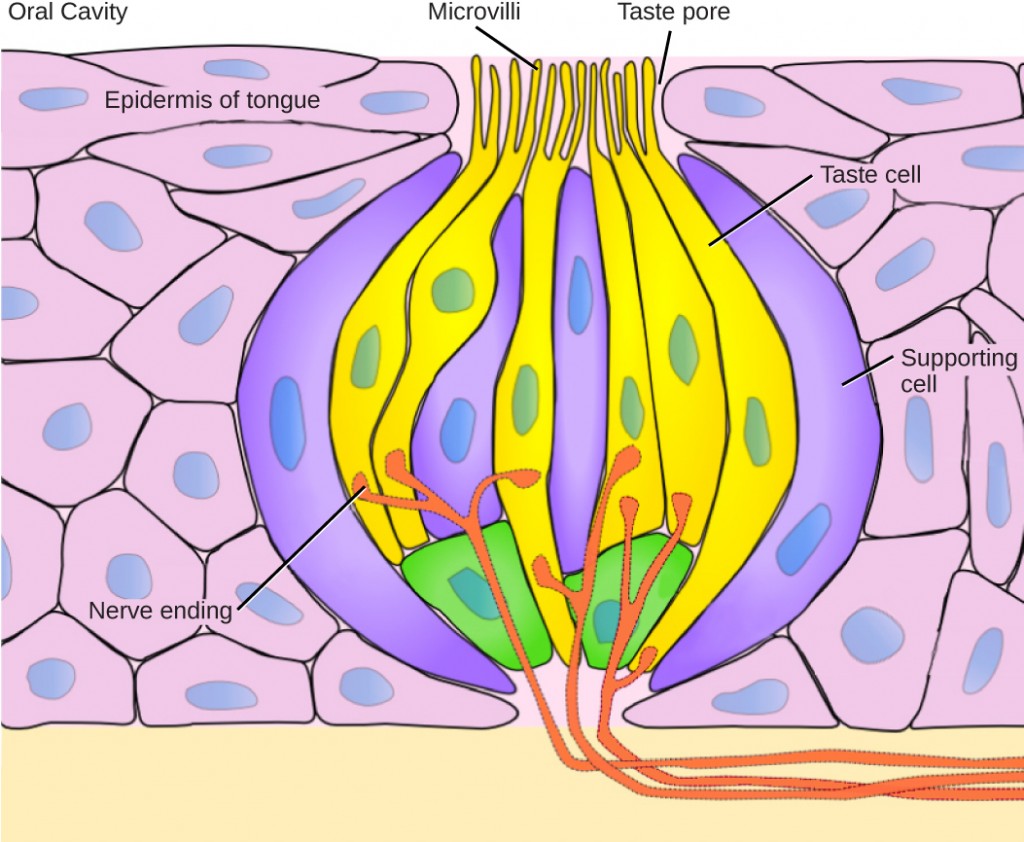
In humans, there are five primary tastes, and each taste has only one corresponding type of receptor. Thus, like olfaction, each receptor is specific to its stimulus (tastant). Transduction of the five tastes happens through different mechanisms that reflect the molecular composition of the tastant. A salty tastant (containing NaCl) provides the sodium ions (Na+) that enter the taste neurons and excite them directly. Sour tastants are acids and belong to the thermoreceptor protein family. Binding of an acid or other sour-tasting molecule triggers a change in the ion channel and these increase hydrogen ion (H+) concentrations in the taste neurons, thus depolarizing them. Sweet, bitter, and umami tastants require a G-protein coupled receptor. These tastants bind to their respective receptors, thereby exciting the specialized neurons associated with them. Both tasting abilities and sense of smell change with age. In humans, the senses decline dramatically by age 50 and continue to decline. A child may find a food to be too spicy, whereas an elderly person may find the same food to be bland and unappetizing.
Smell and taste in the brain
Olfactory neurons project from the olfactory epithelium to the olfactory bulb as thin, unmyelinated axons. The olfactory bulb is composed of neural clusters called glomeruli, and each glomerulus receives signals from one type of olfactory receptor, so each glomerulus is specific to one odorant. From glomeruli, olfactory signals travel directly to the olfactory cortex and then to the frontal cortex and the thalamus. Recall that this is a different path from most other sensory information, which is sent directly to the thalamus before ending up in the cortex. Olfactory signals also travel directly to the amygdala, thereafter reaching the hypothalamus, thalamus, and frontal cortex. The last structure that olfactory signals directly travel to is a cortical center in the temporal lobe structure important in spatial, autobiographical, declarative, and episodic memories. Olfaction is finally processed by areas of the brain that deal with memory, emotions, reproduction, and thought.
Taste neurons project from taste cells in the tongue, esophagus, and palate to the medulla, in the brainstem. From the medulla, taste signals travel to the thalamus and then to the primary gustatory cortex. Information from different regions of the tongue is segregated in the medulla, thalamus, and cortex.
Salty foods activate the taste cells by ______.
a. exciting the taste cell directly
b. causing hydrogen ions to enter the cell
d. causing sodium channels to close
e. binding directly to the receptors
All sensory signals except _____ travel to the _____ in the brain before the cerebral cortex.
a. vision; thalamus
b. olfaction; thalmus
c. vision; cranial nerves
d. olfaction; cranial nerves
From the perspective of the recipient of the signal, in what ways do pheromones differ from other odorants?
What might be the effect on an animal of not being able to perceive taste?
Hearing and vestibular information
Audition, or hearing, is important to humans and to other animals for many different interactions. It enables an organism to detect and receive information about danger, such as an approaching predator, and to participate in communal exchanges like those concerning territories or mating. On the other hand, although it is physically linked to the auditory system, the vestibular system is not involved in hearing. Instead, an animal’s vestibular system detects its own movement, both linear and angular acceleration and deceleration, and balance.
Sound
Auditory stimuli are sound waves, which are mechanical, pressure waves that move through a medium, such as air or water. There are no sound waves in a vacuum since there are no air molecules to move in waves. The speed of sound waves differs, based on altitude, temperature, and medium, but at sea level and a temperature of 20º C (68º F), sound waves travel in the air at about 343 meters per second.
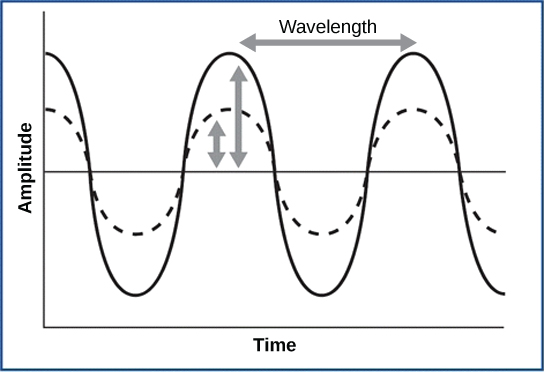
As is true for all waves, there are four main characteristics of a sound wave: frequency, wavelength, period, and amplitude. Frequency is the number of waves per unit of time, and in sound is heard as pitch. High-frequency (≥15.000Hz) sounds are higher-pitched (short wavelength) than low-frequency (long wavelengths; ≤100Hz) sounds. Frequency is measured in cycles per second, and for sound, the most commonly used unit is hertz (Hz), or cycles per second. Most humans can perceive sounds with frequencies between 30 and 20,000 Hz. Women are typically better at hearing high frequencies, but everyone’s ability to hear high frequencies decreases with age. Dogs detect up to about 40,000 Hz; cats, 60,000 Hz; bats, 100,000 Hz; and dolphins 150,000 Hz, and American shad (Alosa sapidissima), a fish, can hear 180,000 Hz. Those frequencies above the human range are called ultrasound.
Amplitude, or the dimension of a wave from peak to trough, in sound is heard as volume and is illustrated in Figure 8.21. The sound waves of louder sounds have greater amplitude than those of softer sounds. For sound, volume is measured in decibels (dB). The softest sound that a human can hear is the zero point. Humans speak normally at 60 decibels.
Reception of sound
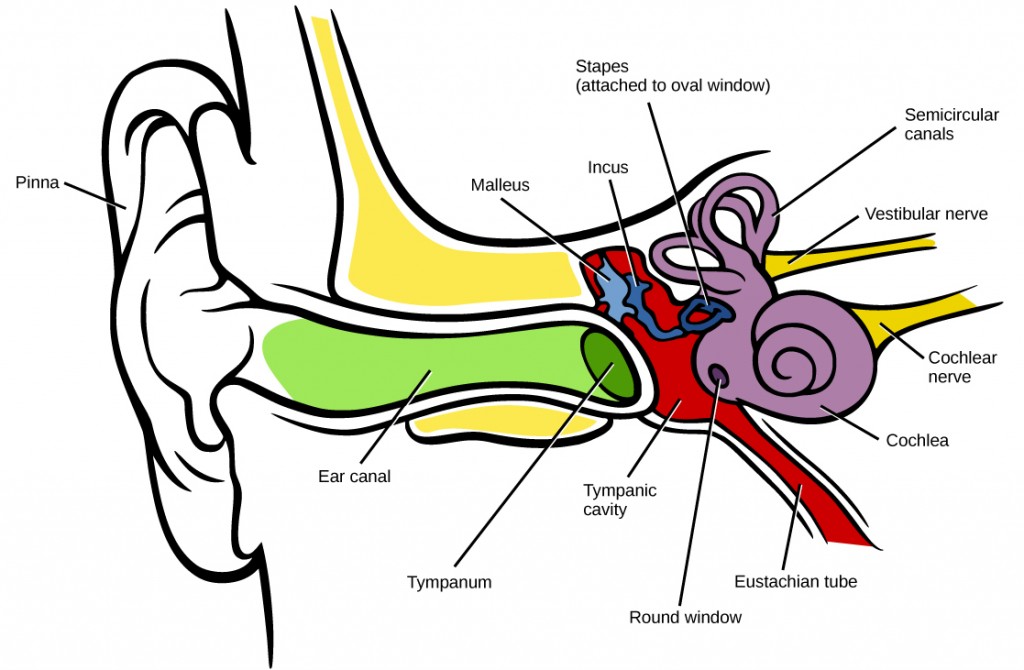
In mammals, sound waves are collected by the external, cartilaginous part of the ear called the pinna, then travel through the auditory canal and cause vibration of the thin diaphragm called the tympanum or ear drum, the innermost part of the outer ear (illustrated in Figure 8.22). Interior to the tympanum is the middle ear. The middle ear holds three small bones called the ossicles, which transfer energy from the moving tympanum to the inner ear. The three ossicles are the malleus(also known as the hammer), the incus (the anvil), and stapes (the stirrup). The aptly named stapes looks very much like a stirrup. The three ossicles are unique to mammals, and each plays a role in hearing. The malleus attaches at three points to the interior surface of the tympanic membrane. The incus attaches the malleus to the stapes. In humans, the stapes is not long enough to reach the tympanum. If we did not have the malleus and the incus, then the vibrations of the tympanum would never reach the inner ear. These bones also function to collect force and amplify sounds. The ear ossicles are homologous to bones in a fish mouth: the bones that support gills in fish are thought to be adapted for use in the vertebrate ear over evolutionary time. Many animals (frogs, reptiles, and birds, for example) use the stapes of the middle ear to transmit vibrations to the middle ear.
Transduction of Sound
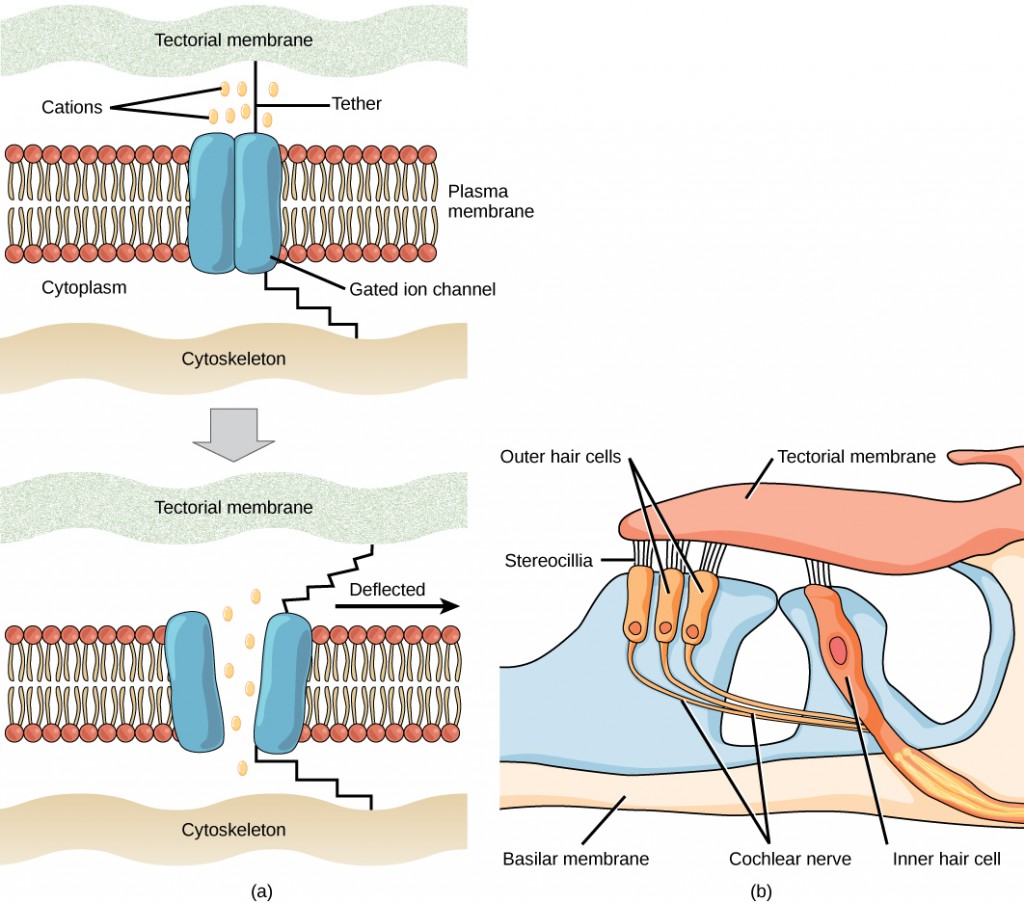
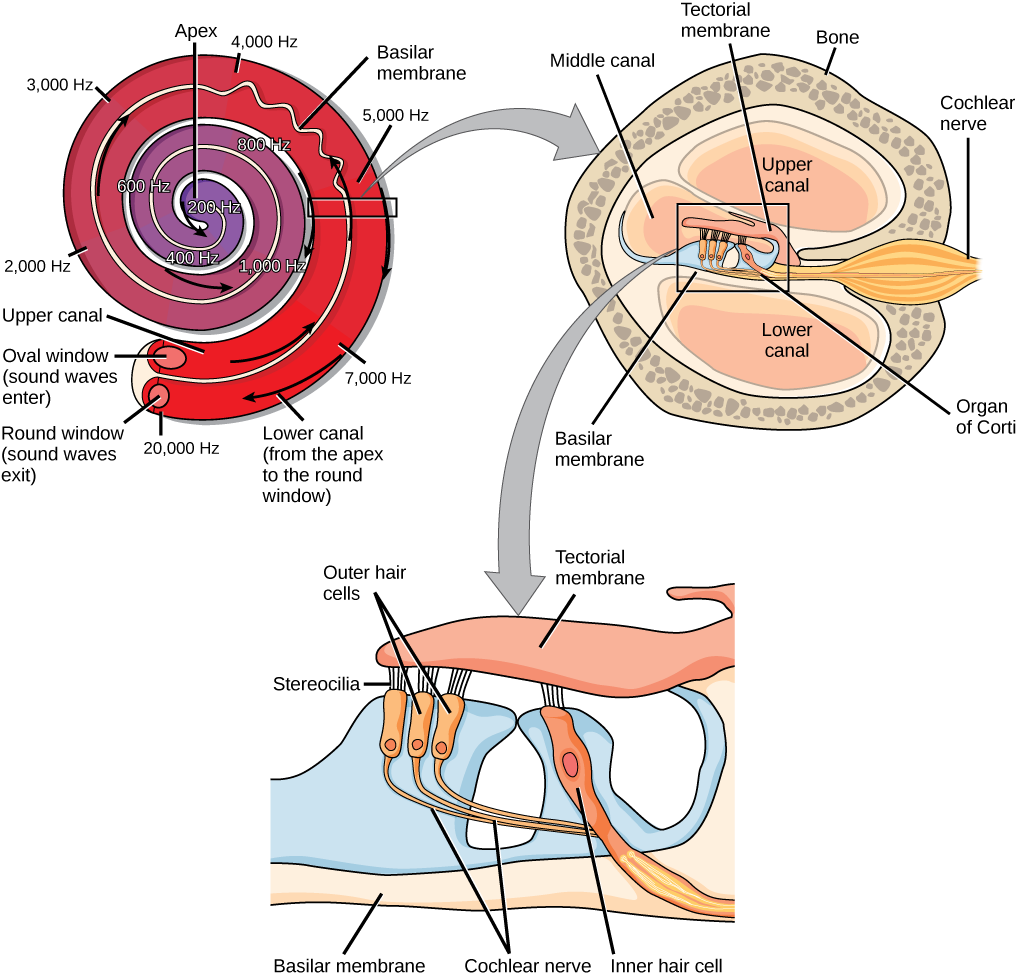
Vibrating objects, such as vocal cords, create sound waves or pressure waves in the air. When these pressure waves reach the ear, the ear transduces this mechanical stimulus (pressure wave) into a nerve impulse (electrical signal) that the brain perceives as sound. The pressure waves strike the tympanum, causing it to vibrate. The mechanical energy from the moving tympanum transmits the vibrations to the three bones of the middle ear. The stapes transmits the vibrations to a thin diaphragm called the oval window, which is the outermost structure of the inner ear. The structures of the inner ear are found in the labyrinth, a bony, hollow structure that is the most interior portion of the ear. Here, the energy from the sound wave is transferred from the stapes through the flexible oval window and to the fluid of the cochlea. The vibrations of the oval window create pressure waves in the fluid (perilymph) inside the cochlea. The cochlea is a whorled structure, like the shell of a snail, and it contains receptors for transduction of the mechanical wave into an electrical signal (as illustrated in Figure 8.23). Inside the cochlea, the basilar membraneis a mechanical analyzer that runs the length of the cochlea, curling toward the cochlea’s center.
The mechanical properties of the basilar membrane change along its length, such that it is thicker, tauter, and narrower at the outside of the whorl (where the cochlea is largest), and thinner, floppier, and broader toward the apex, or center, of the whorl (where the cochlea is smallest). Different regions of the basilar membrane vibrate according to the frequency of the sound wave conducted through the fluid in the cochlea. For these reasons, the fluid-filled cochlea detects different wave frequencies (pitches) at different regions of the membrane. When the sound waves in the cochlear fluid contact the basilar membrane, it flexes back and forth in a wave-like fashion. Above the basilar membrane is the tectorial membrane.
Cochlear implants can restore hearing in people who have a nonfunctional cochlear. The implant consists of a microphone that picks up sound. A speech processor selects sounds in the range of human speech, and a transmitter converts these sounds to electrical impulses, which are then sent to the auditory nerve. Which of the following types of hearing loss would not be restored by a cochlear implant?
a. Hearing loss resulting from absence or loss of hair cells in the organ of Corti.
b. Hearing loss resulting from an abnormal auditory nerve.
c. Hearing loss resulting from fracture of the cochlea.
d. Hearing loss resulting from damage to bones of the middle ear.
The site of transduction is in the organ of Corti (spiral organ). It is composed of hair cells held in place above the basilar membrane like flowers projecting up from soil, with their exposed short, hair-like stereocilia contacting or embedded in the tectorial membrane above them. The inner hair cells are the primary auditory receptors and exist in a single row, numbering approximately 3,500. The stereocilia from inner hair cells extend into small dimples on the tectorial membrane’s lower surface. The outer hair cells are arranged in three or four rows. They number approximately 12,000, and they function to fine tune incoming sound waves. The longer stereocilia that project from the outer hair cells actually attach to the tectorial membrane. All of the stereocilia are mechanoreceptors, and when bent by vibrations they respond by opening a gated ion channel (refer to Figure 8.22). As a result, the hair cell membrane is depolarized, and a signal is transmitted to the chochlear nerve. Intensity (volume) of sound is determined by how many hair cells at a particular location are stimulated.
The hair cells are arranged on the basilar membrane in an orderly way. The basilar membrane vibrates in different regions, according to the frequency of the sound waves impinging on it. Likewise, the hair cells that lay above it are most sensitive to a specific frequency of sound waves. Hair cells can respond to a small range of similar frequencies, but they require stimulation of greater intensity to fire at frequencies outside of their optimal range. The difference in response frequency between adjacent inner hair cells is about 0.2 percent. Compare that to adjacent piano strings, which are about six percent different. Place theory, which is the model for how biologists think pitch detection works in the human ear, states that high frequency sounds selectively vibrate the basilar membrane of the inner ear near the entrance port (the oval window). Lower frequencies travel farther along the membrane before causing appreciable excitation of the membrane. The basic pitch-determining mechanism is based on the location along the membrane where the hair cells are stimulated. The place theory is the first step toward an understanding of pitch perception. Considering the extreme pitch sensitivity of the human ear, it is thought that there must be some auditory “sharpening” mechanism to enhance the pitch resolution.
When sound waves produce fluid waves inside the cochlea, the basilar membrane flexes, bending the stereocilia that attach to the tectorial membrane. Their bending results in action potentials in the hair cells, and auditory information travels along the neural endings of the bipolar neurons of the hair cells (collectively, the auditory nerve) to the brain. When the hairs bend, they release an excitatory neurotransmitter at a synapse with a sensory neuron, which then conducts action potentials to the central nervous system. The cochlear branch of the vestibulocochlear cranial nerve sends information on hearing. The auditory system is very refined, and there is some modulation or “sharpening” built in. The brain can send signals back to the cochlea, resulting in a change of length in the outer hair cells, sharpening or dampening the hair cells’ response to certain frequencies.
Higher processing
The inner hair cells are most important for conveying auditory information to the brain. About 90 percent of the afferent neurons carry information from inner hair cells, with each hair cell synapsing with 10 or so neurons. Outer hair cells connect to only 10 percent of the afferent neurons, and each afferent neuron innervates many hair cells. The afferent, bipolar neurons that convey auditory information travel from the cochlea to the medulla, through the pons and midbrain in the brainstem, finally reaching the primary auditory cortex in the temporal lobe.
Vestibular information
The stimuli associated with the vestibular system are linear acceleration (gravity) and angular acceleration and deceleration. Gravity, acceleration, and deceleration are detected by evaluating the inertia on receptive cells in the vestibular system. Gravity is detected through head position. Angular acceleration and deceleration are expressed through turning or tilting of the head.
The vestibular system has some similarities with the auditory system. It utilizes hair cells just like the auditory system, but it excites them in different ways. There are five vestibular receptor organs in the inner ear: the utricle, the saccule, and three semicircular canals. Together, they make up what’s known as the vestibular labyrinth that is shown in Figure 8.24. The utricle and saccule respond to acceleration in a straight line, such as gravity. The roughly 30,000 hair cells in the utricle and 16,000 hair cells in the saccule lie below a gelatinous layer, with their stereocilia projecting into the gelatin. Embedded in this gelatin are calcium carbonate crystals—like tiny rocks. When the head is tilted, the crystals continue to be pulled straight down by gravity, but the new angle of the head causes the gelatin to shift, thereby bending the stereocilia. The bending of the stereocilia stimulates the neurons, and they signal to the brain that the head is tilted, allowing the maintenance of balance. It is the vestibular branch of the vestibulocochlear cranial nerve that deals with balance.
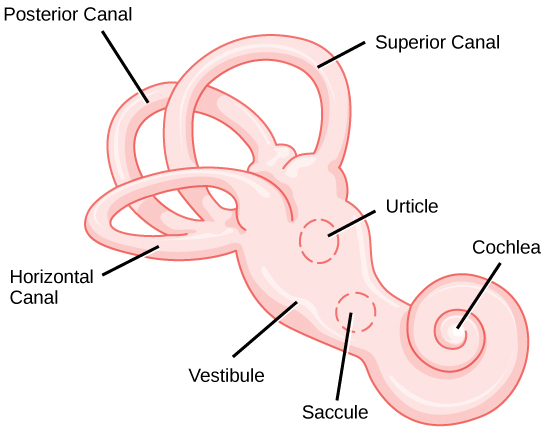
The fluid-filled semicircular canals are tubular loops set at oblique angles. They are arranged in three spatial planes. The base of each canal has a swelling that contains a cluster of hair cells. The hairs project into a gelatinous cap called the cupula and monitor angular acceleration and deceleration from rotation. They would be stimulated by driving your car around a corner, turning your head, or falling forward. One canal lies horizontally, while the other two lie at about 45 degree angles to the horizontal axis, as illustrated in Figure 8.24. When the brain processes input from all three canals together, it can detect angular acceleration or deceleration in three dimensions. When the head turns, the fluid in the canals shifts, thereby bending stereocilia and sending signals to the brain. Upon cessation accelerating or decelerating—or just moving—the movement of the fluid within the canals slows or stops. For example, imagine holding a glass of water. When moving forward, water may splash backwards onto the hand, and when motion has stopped, water may splash forward onto the fingers. While in motion, the water settles in the glass and does not splash. Note that the canals are not sensitive to velocity itself, but to changes in velocity, so moving forward at 60mph with your eyes closed would not give the sensation of movement, but suddenly accelerating or braking would stimulate the receptors.
Hair cells from the utricle, saccule, and semicircular canals also communicate through bipolar neurons to the cochlear nucleus in the medulla. Cochlear neurons send descending projections to the spinal cord and ascending projections to the pons, thalamus, and cerebellum. Connections to the cerebellum are important for coordinated movements. There are also projections to the temporal cortex, which account for feelings of dizziness; projections to autonomic nervous system areas in the brainstem, which account for motion sickness; and projections to the primary somatosensory cortex, which monitors subjective measurements of the external world and self-movement. People with lesions in the vestibular area of the somatosensory cortex see vertical objects in the world as being tilted. Finally, the vestibular signals project to certain optic muscles to coordinate eye and head movements.
Auditory hair cells are indirectly anchored to the _____.
a. basilar membrane
b. oval window
c. tectorial membrane
d. ossicles
Which of the following are found both in the auditory system and the vestibular system?
a. basilar membrane
b. hair cells
c. semicircular canals
d. ossicles
How would a rise in altitude likely affect the speed of a sound transmitted through air?
How might being in a place with less gravity than Earth has (such as Earth’s moon) affect vestibular sensation, and why?
Vision
Vision is the ability to detect light patterns from the outside environment and interpret them into images. Animals are bombarded with sensory information, and the sheer volume of visual information can be problematic. Fortunately, the visual systems of species have evolved to attend to the most-important stimuli. The importance of vision to humans is further substantiated by the fact that about one-third of the human cerebral cortex is dedicated to analyzing and perceiving visual information.
Light
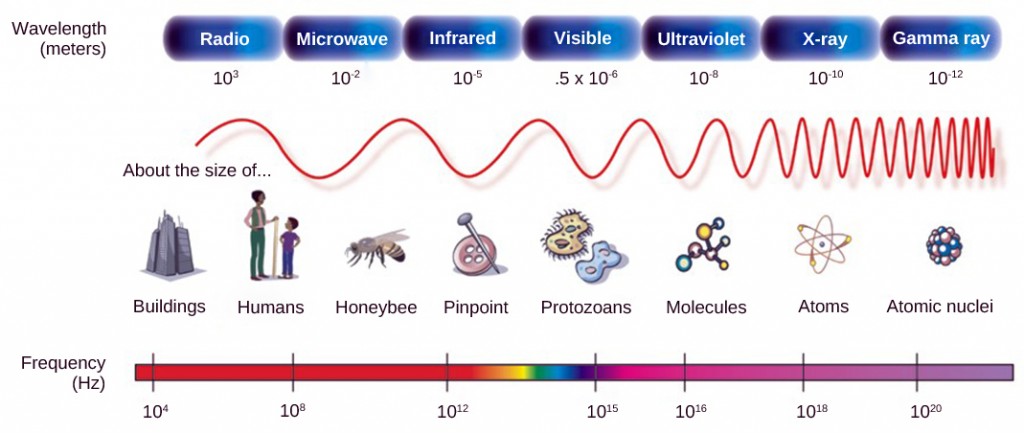
As with auditory stimuli, light travels in waves. The compression waves that compose sound must travel in a medium—a gas, a liquid, or a solid. In contrast, light is composed of electromagnetic waves and needs no medium; light can travel in a vacuum (Figure 8.25). The behavior of light can be discussed in terms of the behavior of waves and also in terms of the behavior of the fundamental unit of light—a packet of electromagnetic radiation called a photon. A glance at the electromagnetic spectrum shows that visible light for humans is just a small slice of the entire spectrum, which includes radiation that we cannot see as light because it is below the frequency of visible red light and above the frequency of visible violet light.
Certain variables are important when discussing perception of light. Wavelength (which varies inversely with frequency) manifests itself as hue. Light at the red end of the visible spectrum has longer wavelengths (and is lower frequency), while light at the violet end has shorter wavelengths (and is higher frequency). The wavelength of light is expressed in nanometers (nm); one nanometer is one billionth of a meter. Humans perceive light that ranges between approximately 380 nm and 740 nm. Some other animals, though, can detect wavelengths outside of the human range. For example, bees see near-ultraviolet light in order to locate nectar guides on flowers, and some non-avian reptiles sense infrared light (heat that prey gives off).
Wave amplitude is perceived as luminous intensity, or brightness. The standard unit of intensity of light is the candela, which is approximately the luminous intensity of a one common candle.
Light waves travel 299,792 km per second in a vacuum, (and somewhat slower in various media such as air and water), and those waves arrive at the eye as long (red), medium (green), and short (blue) waves. What is termed “white light” is light that is perceived as white by the human eye. This effect is produced by light that stimulates equally the color receptors in the human eye. The apparent color of an object is the color (or colors) that the object reflects. Thus a red object reflects the red wavelengths in mixed (white) light and absorbs all other wavelengths of light.
Anatomy of the eye
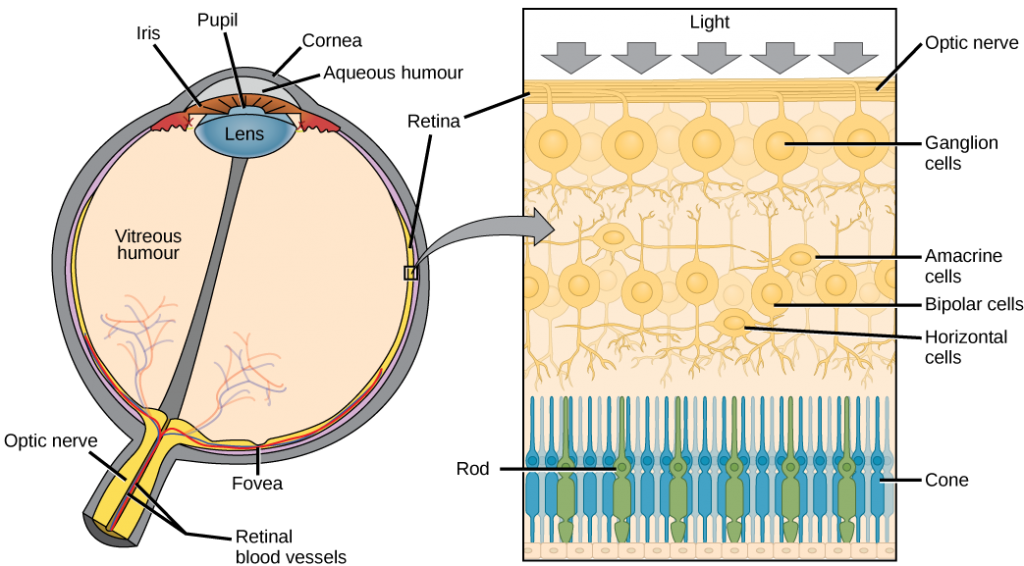
The photoreceptive cells of the eye, where transduction of light to nervous impulses occurs, are located in the retina (Figure 8.26) on the inner surface of the back of the eye. But light does not impinge on the retina unaltered. It passes through other layers that process it so that it can be interpreted by the retina (Figure 8.25b). The cornea, the front transparent layer of the eye, and the crystalline lens, a transparent convex structure behind the cornea, both refract (bend) light to focus the image on the retina. The iris, which is conspicuous as the colored part of the eye, is a circular muscular ring lying between the lens and cornea that regulates the amount of light entering the eye. In conditions of high ambient light, the iris contracts, reducing the size of the pupil at its center. In conditions of low light, the iris relaxes and the pupil enlarges.
Which of the following statements about the human eye is false?
a. Rods detect color, while cones detect only shades of gray.
b. When light enters the retina, it passes the ganglion cells and bipolar cells before reaching photoreceptors at the rear of the eye.
c. The iris adjusts the amount of light coming into the eye.
d. The cornea is a protective layer on the front of the eye.
There are two types of photoreceptors in the retina: rods and cones, named for their general appearance as illustrated in Figure 8.27. Rods are strongly photosensitive and are located in the outer edges of the retina. They detect dim light and are used primarily for peripheral and nighttime vision. Cones are weakly photosensitive and are located near the center of the retina. They respond to bright light, and their primary role is in daytime, color vision.
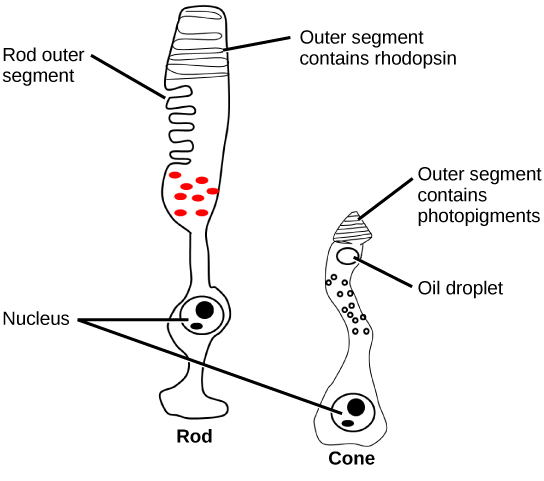
Figure 8.27. Rods and cones are photoreceptors in the retina. Rods respond in low light and can detect only shades of gray. Cones respond in intense light and are responsible for color vision. (credit: modification of work by Piotr Sliwa)
The fovea is the region in the center back of the eye that is responsible for acute vision. The fovea has a high density of cones. When you bring your gaze to an object to examine it intently in bright light, the eyes orient so that the object’s image falls on the fovea. However, when looking at a star in the night sky or other object in dim light, the object can be better viewed by the peripheral vision because it is the rods at the edges of the retina, rather than the cones at the center, that operate better in low light. In humans, cones far outnumber rods in the fovea.
Transduction of light
The rods and cones are the site of transduction of light to a neural signal. Both rods and cones contain photopigments. In vertebrates, the main photopigment, rhodopsin, has two main parts: an opsin, which is a membrane protein (in the form of a cluster of α-helices that span the membrane), and retinal—a molecule that absorbs light. When light hits a photoreceptor, it causes a shape change in the retinal, altering its structure from a bent (cis) form of the molecule to its linear (trans) isomer. This isomerization of retinal activates the rhodopsin, starting a cascade of events that ends with the closing of Na+ channels in the membrane of the photoreceptor. Thus, unlike most other sensory neurons (which become depolarized by exposure to a stimulus) visual receptors become hyperpolarized and thus driven away from threshold (Figure 8.28).
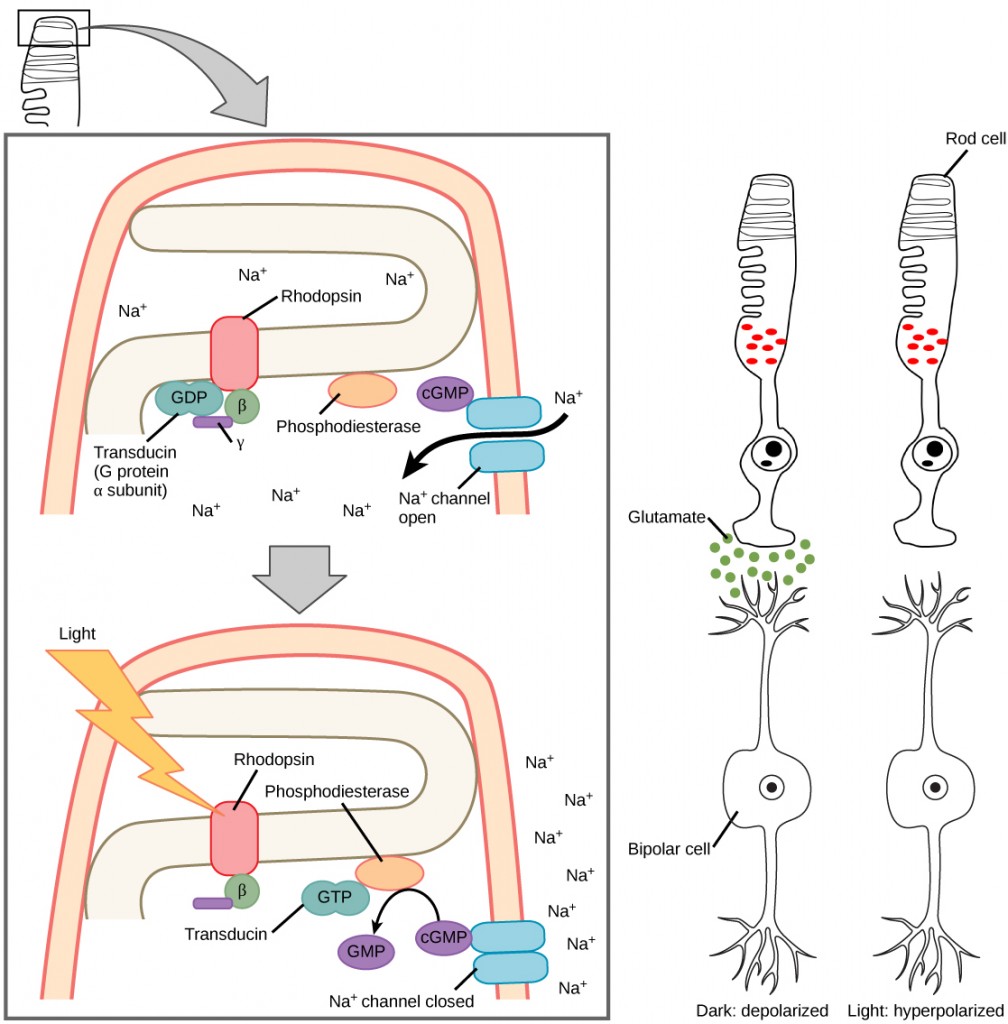
Higher processing
The myelinated axons of ganglion cells make up the optic nerves. Within the nerves, different axons carry different qualities of the visual signal. Some axons constitute the magnocellular (big cell) pathway, which carries information about form, movement, depth, and differences in brightness. Other axons constitute the parvocellular (small cell) pathway, which carries information on color and fine detail. Some visual information projects directly back into the brain, while other information crosses to the opposite side of the brain. This crossing of optical pathways produces the distinctive optic chiasma (Greek, for “crossing”) found at the base of the brain and allows us to coordinate information from both eyes.
Once in the brain, visual information is processed in several places, and its routes reflect the complexity and importance of visual information to humans and other animals. One route takes the signals to the thalamus, which serves as the routing station for all incoming sensory impulses except olfaction. In the thalamus, the magnocellular and parvocellular distinctions remain intact, and there are different layers of the thalamus dedicated to each. When visual signals leave the thalamus, they travel to the primary visual cortex at the rear of the brain. From the visual cortex, the visual signals travel in two directions. One stream that projects to the parietal lobe, in the side of the brain, carries magnocellular (“where”) information. A second stream projects to the temporal lobe and carries both magnocellular (“where”) and parvocellular (“what”) information.
Another important visual route is a pathway from the retina to the superior colliculus in the midbrain, where eye movements are coordinated and integrated with auditory information. Finally, there is the pathway from the retina to the suprachiasmatic nucleus (SCN) of the hypothalamus. The SCN is a cluster of cells that is considered to be the body’s internal clock, which controls our circadian (day-long) cycle. The SCN sends information to the pineal gland, which is important in sleep/wake patterns and annual cycles.
How could the pineal gland, the brain structure that plays a role in annual cycles, use visual information from the suprachiasmatic nucleus of the hypothalamus?
Why is it easier to see images at night using peripheral, rather than the central, vision?
a. Cones are denser in the periphery of the retina.
b. Bipolar cells are denser in the periphery of the retina.
c. Rods are denser in the periphery of the retina.
d. The optic nerve exits at the periphery of the retina.
What sensory information do animals use that human cannot? Provide three different examples and briefly describe each.