
4.2: Ion and Water Balance on Land
- Page ID
- 9556

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Explain how ion and water balance function in sample animal terrestrial systems.
Although the kidneys are the major osmoregulatory organ, the skin and lungs also play a role in the process. Water and electrolytes are lost through sweat glands in the skin, which helps moisturize and cool the skin surface, while the lungs expel a small amount of water in the form of mucous secretions and via evaporation of water vapor.
Kidneys: the main osmoregulatory organ
The kidneys, illustrated in Figure 4.4, are a pair of bean-shaped structures that are located just below and posterior to the liver in the peritoneal cavity. The adrenal glands sit on top of each kidney and are also called the suprarenal glands. Kidneys filter blood and purify it. All the blood in the human body is filtered many times a day by the kidneys; these organs use up almost 25 percent of the oxygen absorbed through the lungs to perform this function. Oxygen allows the kidney cells to efficiently manufacture chemical energy in the form of ATP through aerobic respiration. The filtrate coming out of the kidneys is called urine.

Kidney structure
Externally, the kidneys are surrounded by three layers, illustrated in Figure 4.5. The outermost layer is a tough connective tissue layer called the renal fascia. The second layer is called the perirenal fat capsule, which helps anchor the kidneys in place. The third and innermost layer is the renal capsule. Internally, the kidney has three regions—an outer cortex, a medulla in the middle, and the renal pelvis in the region called the hilum of the kidney. The hilum is the concave part of the bean-shape where blood vessels and nerves enter and exit the kidney; it is also the point of exit for the ureters. The renal cortex is granular due to the presence of nephrons—the functional unit of the kidney. The medulla consists of multiple pyramidal tissue masses, called the renal pyramids. In between the pyramids are spaces called renal columns through which the blood vessels pass. The tips of the pyramids, called renal papillae, point toward the renal pelvis. There are, on average, eight renal pyramids in each kidney. The renal pyramids along with the adjoining cortical region are called the lobes of the kidney. The renal pelvis leads to the ureter on the outside of the kidney. On the inside of the kidney, the renal pelvis branches out into two or three extensions called the major calyces, which further branch into the minor calyces. The ureters are urine-bearing tubes that exit the kidney and empty into the urinary bladder.

Which of the following statements about the kidney is false?
a. The renal pelvis drains into the ureter.
b. The renal pyramids are in the medulla.
c. The cortex covers the capsule.
d. Nephrons are in the renal cortex
Because the kidney filters blood, its network of blood vessels is an important component of its structure and function. The arteries, veins, and nerves that supply the kidney enter and exit at the renal hilum. Renal blood supply starts with the branching of the aorta into the renal arteries(which are each named based on the region of the kidney they pass through) and ends with the exiting of the renal veins to join the inferior vena cava. The renal arteries split into several segmental arteries upon entering the kidneys. Each segmental artery splits further into several interlobar arteries and enters the renal columns, which supply the renal lobes. The interlobar arteries split at the junction of the renal cortex and medulla to form the arcuate arteries. The arcuate “bow-shaped” arteries form arcs along the base of the medullary pyramids. Corticalradiate arteries, as the name suggests, radiate out from the arcuate arteries. The cortical radiate arteries branch into numerous afferent arterioles and then enter the capillaries supplying the nephrons. Veins trace the path of the arteries and have similar names, except there are no segmental veins.
As mentioned previously, the functional unit of the kidney is the nephron, illustrated in Figure 4.6. Each kidney is made up of over one million nephrons that dot the renal cortex, giving it a granular appearance when sectioned sagittally. There are two types of nephrons— cortical nephrons (85 percent), which are deep in the renal cortex, and juxtamedullary nephrons, which lie in the renal cortex close to the renal medulla. A nephron consists of three parts—a renal corpuscle, a renal tubule, and the associated capillary network, which originates from the cortical radiate arteries.

Nephrons: the functional unit
Nephrons take a simple filtrate of the blood and modify it into the urine. Many changes take place in the different parts of the nephron before the urine is created for disposal. The term forming urine will be used hereafter to describe the filtrate as it is modified into the true urine. The principal task of the nephron population is to balance the plasma to homeostatic set points and excrete potential toxins in the urine. They do this by accomplishing three principle functions—filtration, reabsorption, and secretion. They also have additional secondary functions that exert control in three areas: blood pressure (via production of renin), red blood cell production (via the hormone EPO), and calcium absorption (via conversion of calcidiol into calcitriol, the active form of vitamin D).
Renal Corpuscle
The renal corpuscle, located in the renal cortex, is made up of a network of capillaries known as the glomerulus and the capsule, a cup-shaped chamber that surrounds it, called the glomerular or Bowman’s capsule. The renal corpuscle consists of a tuft of capillaries called the glomerulus that is largely surrounded by Bowman’s (glomerular) capsule. The glomerulus is a high-pressure capillary bed between afferent and efferent arterioles. Bowman’s capsule surrounds the glomerulus to form a lumen and captures and directs this filtrate to the PCT. The outermost part of Bowman’s capsule, the parietal layer, is a simple squamous epithelium. It transitions onto the glomerular capillaries in an intimate embrace to form the visceral layer of the capsule. Here, the cells are not squamous, but uniquely shaped cells (podocytes) extending finger-like arms (pedicels) to cover the glomerular capillaries (Figure 4.7). These projections interdigitate to form filtration slits, leaving small gaps between the digits to form a sieve. As blood passes through the glomerulus, 10 to 20 percent of the plasma filters between these sieve-like fingers to be captured by Bowman’s capsule and funneled to the PCT. Where the fenestrae (windows) in the glomerular capillaries match the spaces between the podocyte “fingers,” the only thing separating the capillary lumen and the lumen of Bowman’s capsule is their shared basement membrane (Figure 4.8). These three features comprise what is known as the filtration membrane. This membrane permits very rapid movement of filtrate from capillary to capsule through pores that are only 70 nm in diameter.


The fenestrations prevent filtration of blood cells or large proteins but allow most other constituents through. These substances cross readily if they are less than 4 nm in size and most pass freely up to 8 nm in size. An additional factor affecting the ability of substances to cross this barrier is their electric charge. The proteins associated with these pores are negatively charged, so they tend to repel negatively charged substances and allow positively charged substances to pass more readily. The basement membrane prevents filtration of medium-to-large proteins such as globulins. There are also mesangial cells in the filtration membrane that can contract to help regulate the rate of filtration of the glomerulus. Overall, filtration is regulated by fenestrations in capillary endothelial cells, podocytes with filtration slits, membrane charge, and the basement membrane between capillary cells. The result is the creation of a filtrate that does not contain cells or large proteins and has a slight predominance of positively charged substances.
Lying just outside Bowman’s capsule and the glomerulus is the juxtaglomerular apparatus (JGA) (Figure 4.9). At the juncture where the afferent and efferent arterioles enter and leave Bowman’s capsule, the initial part of the distal convoluted tubule (DCT) comes into direct contact with the arterioles. The wall of the DCT at that point forms a part of the JGA known as the macula densa. This cluster of cuboidal epithelial cells monitors the fluid composition of fluid flowing through the DCT. In response to the concentration of Na+ in the fluid flowing past them, these cells release paracrine signals. They also have a single, nonmotile cilium that responds to the rate of fluid movement in the tubule. The paracrine signals released in response to changes in flow rate and Na+concentration are adenosine triphosphate (ATP) and adenosine.
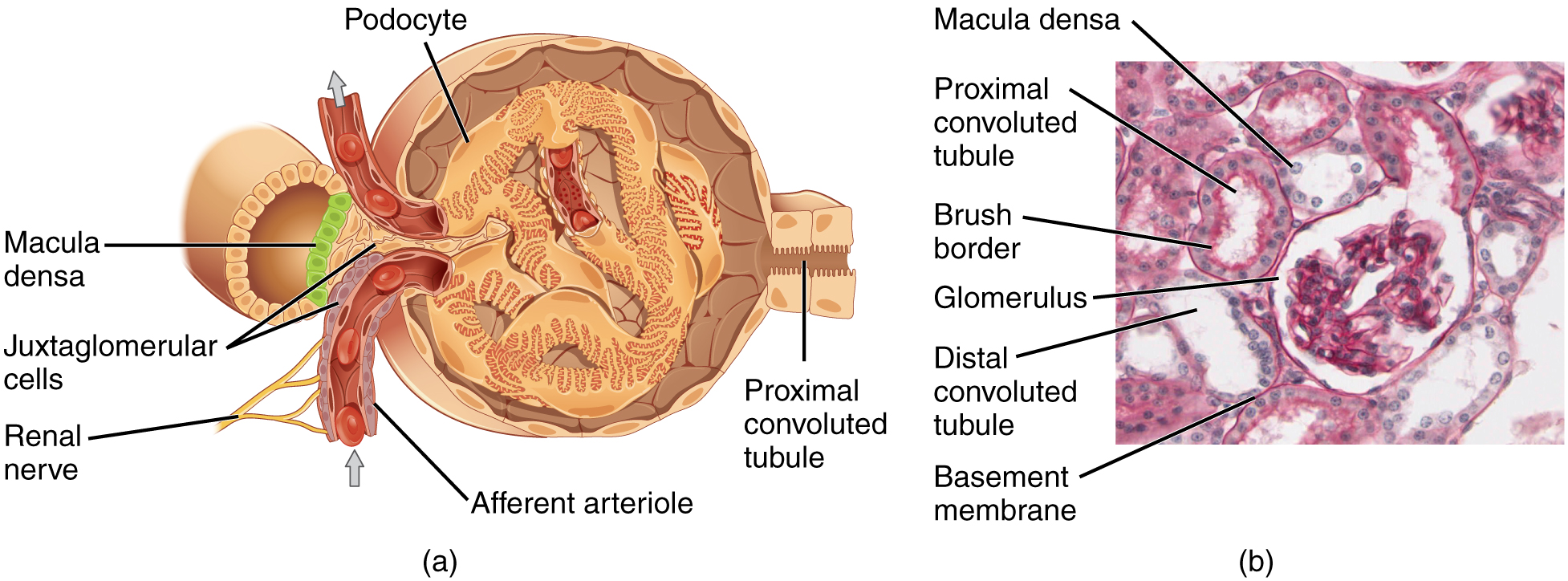
The renal tubule is a long and convoluted structure that emerges from the glomerulus and can be divided into three parts based on function. The first part is called the proximal convoluted tubule (PCT) due to its proximity to the glomerulus; it stays in the renal cortex. The second part is called the loop of Henle, or nephritic loop because it forms a loop (with descending and ascending limbs) that goes through the renal medulla. The third part of the renal tubule is called the distal convoluted tubule (DCT) and this part is also restricted to the renal cortex. The DCT, which is the last part of the nephron, connects and empties its contents into collecting ducts that line the medullary pyramids. The collecting ducts amass contents from multiple nephrons and fuse together as they enter the papillae of the renal medulla.
Capillary network within the nephron
The capillary network that originates from the renal arteries supplies the nephron with blood that needs to be filtered. The branch that enters the glomerulus is called the afferent arteriole. The branch that exits the glomerulus is called the efferent arteriole. Within the glomerulus, the network of capillaries is called the glomerular capillary bed. Once the efferent arteriole exits the glomerulus, it forms the peritubular capillary network, which surrounds and interacts with parts of the renal tubule. In cortical nephrons, the peritubular capillary network surrounds the PCT and DCT. In juxtamedullary nephrons, the peritubular capillary network forms a network around the loop of Henle and is called the vasa recta.
Which of the following statements about the nephron is false?
a. The collecting duct empties into the distal convoluted tubule.
b. The Bowman’s capsule surrounds the glomerulus.
c. The loop of Henle is between the proximal and distal convoluted tubules.
d. The loop of Henle empties into the distal convoluted tubule
Kidney function and physiology
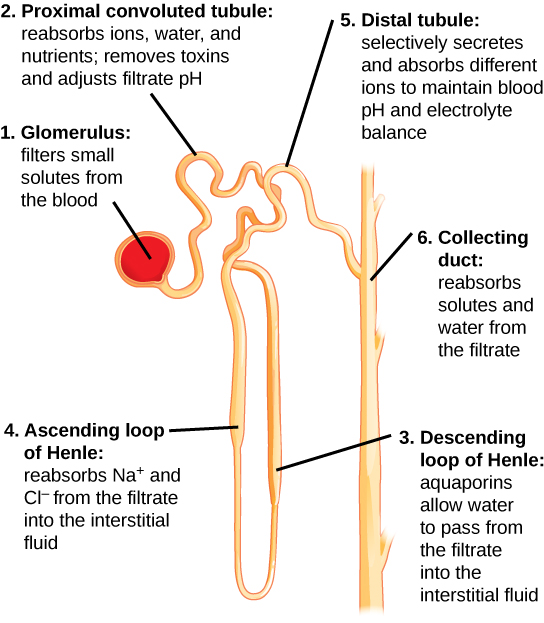
Kidneys filter blood in a three-step process. First, the nephrons filter blood that runs through the capillary network in the glomerulus. Almost all solutes, except for proteins, are filtered out into the glomerulus by a process called glomerular filtration. Second, the filtrate is collected in the renal tubules. Most of the solutes get reabsorbed in the PCT by a process called tubular reabsorption. In the loop of Henle, the filtrate continues to exchange solutes and water with the renal medulla and the peritubular capillary network. Water is also reabsorbed during this step. Then, additional solutes and wastes are secreted into the kidney tubules during tubular secretion, which is, in essence, the opposite process of tubular reabsorption. The collecting ducts collect filtrate coming from the nephrons and fuse in the medullary papillae. From here, the papillae deliver the filtrate, now called urine, into the minor calyces that eventually connect to the ureters through the renal pelvis. This entire process is illustrated in Figure 4.10.
Glomerular filtration
Glomerular filtration filters out most of the solutes due to high blood pressure and specialized membranes in the afferent arteriole. The blood pressure in the glomerulus is maintained independent of factors that affect systemic blood pressure. The “leaky” connections between the endothelial cells of the glomerular capillary network allow solutes to pass through easily. All solutes in the glomerular capillaries, except for macromolecules like proteins, pass through by passive diffusion. There is no energy requirement at this stage of the filtration process. Glomerular filtration rate (GFR) is the volume of glomerular filtrate formed per minute by the kidneys. GFR is regulated by multiple mechanisms and is an important indicator of kidney function.
Tubular reabsorption and secretion
Tubular reabsorption occurs in the PCT part of the renal tubule. Almost all nutrients are reabsorbed, and this occurs either by passive or active transport. Reabsorption of water and some key electrolytes are regulated and can be influenced by hormones. Sodium (Na+) is the most abundant ion and most of it is reabsorbed by active transport and then transported to the peritubular capillaries. Because Na+ is actively transported out of the tubule, water follows it to even out the osmotic pressure. Water is also independently reabsorbed into the peritubular capillaries due to the presence of aquaporins, or water channels, in the PCT. This occurs due to the low blood pressure and high osmotic pressure in the peritubular capillaries. However, every solute has a transport maximum and the excess is not reabsorbed.
In the loop of Henle, the permeability of the membrane changes. The descending limb is permeable to water, not solutes; the opposite is true for the ascending limb. Additionally, the loop of Henle invades the renal medulla, which is naturally high in salt concentration and tends to absorb water from the renal tubule and concentrate the filtrate. The osmotic gradient increases as it moves deeper into the medulla. Because two sides of the loop of Henle perform opposing functions, as illustrated in Figure 4.11, it acts as a countercurrent multiplier. The vasa recta around it acts as the countercurrent exchanger.

Loop diuretics are drugs sometimes used to treat hypertension. These drugs inhibit the reabsorption of Na+ and Cl– ions by the ascending limb of the loop of Henle. A side effect is that they increase urination. Why do you think this is the case?
By the time the filtrate reaches the DCT, most of the urine and solutes have been reabsorbed. If the body requires additional water, all of it can be reabsorbed at this point. Further reabsorption is controlled by hormones, which will be discussed in a later section. Excretion of wastes occurs due to lack of reabsorption combined with tubular secretion. Undesirable products like metabolic wastes, urea, uric acid, and certain drugs, are excreted by tubular secretion. Most of the tubular secretion happens in the DCT, but some occurs in the early part of the collecting duct. Kidneys also maintain an acid-base balance by secreting excess H+ ions.
Although parts of the renal tubules are named proximal and distal, in a cross-section of the kidney, the tubules are placed close together and in contact with each other and the glomerulus. This allows for the exchange of chemical messengers between the different cell types. For example, the DCT ascending limb of the loop of Henle has masses of cells called macula densa, which are in contact with cells of the afferent arterioles called juxtaglomerular cells. Together, the macula densa and juxtaglomerular cells form the juxtaglomerular complex (JGC). The JGC is an endocrine structure that secretes the enzyme renin and the hormone erythropoietin. When hormones trigger the macula densa cells in the DCT due to variations in blood volume, blood pressure, or electrolyte balance, these cells can immediately communicate the problem to the capillaries in the afferent and efferent arterioles, which can constrict or relax to change the glomerular filtration rate of the kidneys.