
9.4: Selenium
- Page ID
- 40978

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Selenium can be divided into 2 categories: organic and inorganic. The organic forms contain carbon, while the inorganic forms do not. The primary inorganic forms of selenium are selenite (\(\ce{SeO3}\)) and selenate (\(\ce{SeO4}\)). Selenite and selenate are not commonly found alone in nature; they are usually complexed with sodium to form sodium selenite (\(\ce{Na2SeO3}\)) and sodium selenate (\(\ce{Na2SeO4}\))1.
Query \(\PageIndex{1}\)
Selenomethionine is the most common organic form of selenium. The structure of selenomethionine is shown above the structure of the amino acid methionine in the figure below.
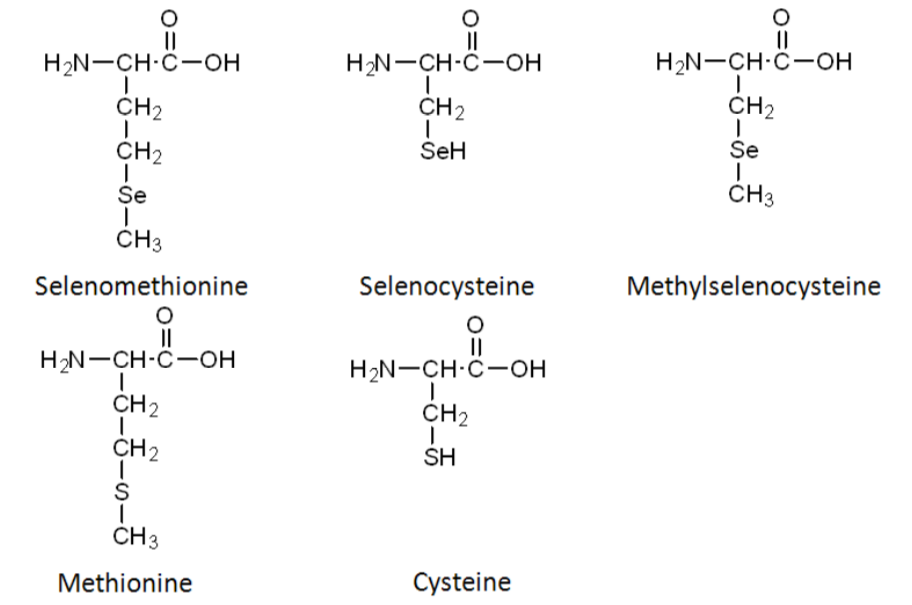
In comparing the structures of selenomethionine or methionine, you can see that the only difference is that selenium has been substituted for sulfur (S) in methionine. Selenocysteine is considered the 21st amino acid by some, because there is a codon that directs its insertion into selenoproteins. Like selenomethionine versus methionine, the only difference between selenocysteine and cysteine is the substitution of selenium for sulfur. The last organic form is methylselenocysteine (aka Se-methylselenocysteine). Notice that its structure is like selenocysteine, but with a methyl group added (like the name suggests).
The selenium content of plants is dependent on the soil where they are grown. As shown below, soil selenium levels vary greatly throughout the United States, meaning that the selenium content of plant foods also vary greatly.
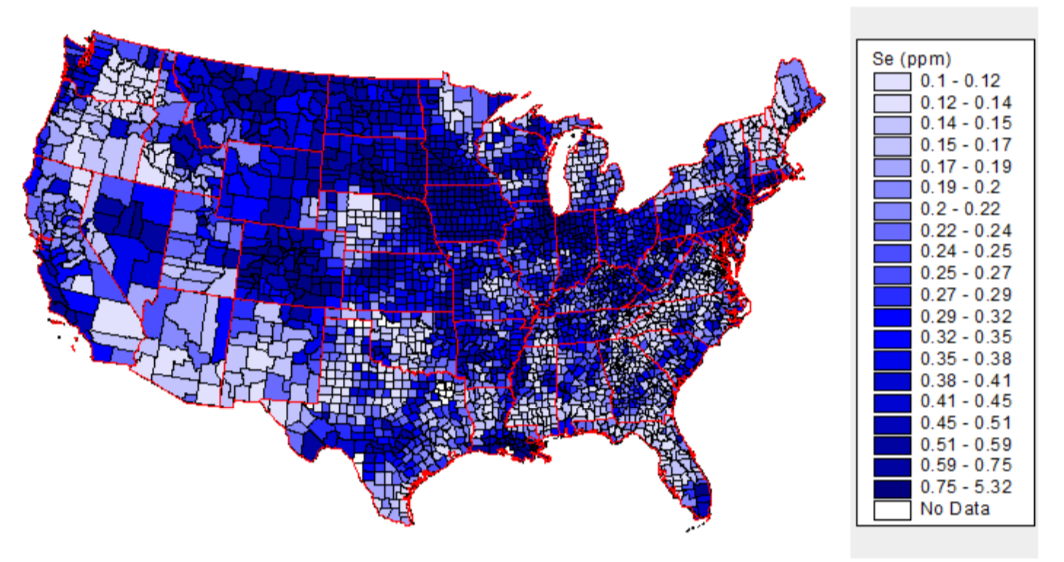
The above map is interactive, so to see the soil selenium levels in a certain county or state, click on it in the link below.
Web Link
Inorganic forms of selenium are commonly used in supplements. Selenomethionine is the most common organic form of selenium in supplements and food. It is found in cereal grains such as wheat, corn, and rice as well as soy. Yeast are typically used to produce selenomethionine for supplements.
It should be noted that selenomethionine accumulates at much higher concentrations in the body than other forms of selenium. This is because it can be nonspecifically incorporated into body proteins in place of methionine. However, despite accumulating at higher levels, selenomethionine is less effective than the methylselenocysteine in decreasing cancer incidence or growth in animal models3. However, methylselenocysteine is not commonly consumed, because it is a form that plants accumulate to prevent selenium from becoming toxic to themselves. Thus, plants need to be grown in the presence of high selenium levels to accumulate meaningful amounts of methylselenocysteine.
Query \(\PageIndex{2}\)
Selenoproteins
As mentioned earlier, selenium's antioxidant function is not due to the mineral itself, but a result of the action of selenoenzymes. Selenoenzymes is a term used to describe the subset of selenoproteins that are enzymes. This is illustrated in the figure below, where the different colored circles represent amino acids in the crescent shaped enzyme. In most enzymes, the mineral is a cofactor that is external to the enzyme, as shown on the left. Selenoenzymes contain selenocysteine as an amino acid in the active site of the enzyme. Thus, in selenoenzymes, selenium does not serve as a cofactor, which is different than most minerals required for enzyme function.
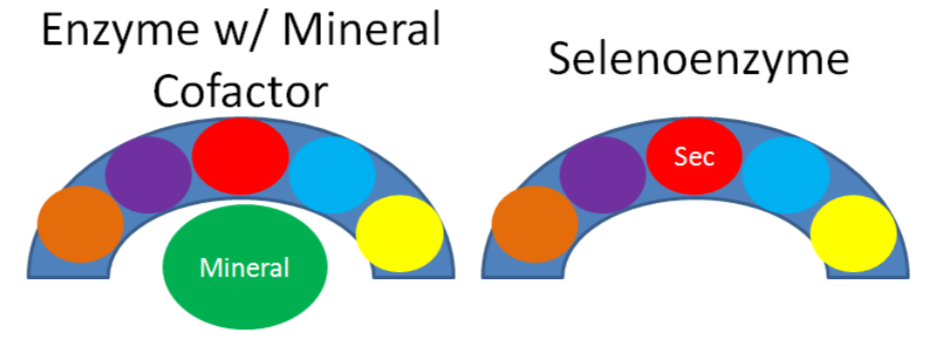
Query \(\PageIndex{3}\)
25 human selenoproteins, containing the amino acid selenocysteine, have been identified. The following table lists these selenoproteins along with their function.
| Selenoprotein | Function |
|---|---|
| Glutathione peroxidase 1 (GPX1) | Cytosolic glutathione peroxidase |
| Glutathione peroxidase 2 (GPX2) | Gastrointestinal glutathione peroxidase |
| Glutathione peroxidase 3 (GPX3) | Plasma glutathione peroxidase |
| Glutathione peroxidase 4 (GPX4) | Phospholipid hydroperoxide glutathione peroxidase |
| Glutathione peroxidase 6 (GPX6) | Olfactory glutathione peroxidase |
| Iodothyronine 5'-deiodinase-1 (DI1) | Thyroid hormone-activating iodothyronine deodinase |
| Iodothyronine 5'-deiodinase-2 (DI2) | Tissue-specific thyroid hormone-activating iodothyronine deodinase |
| Iodothyronine 5'-deiodinase-3 (DI3) | Tissue-specific thyroid hormone-deactivating iodothyronine deodinase |
| Thioredoxin reductase (TR1) | Reduction of cytosolic thioredoxin |
| Thioredoxin reductase (TR2) | Testis-specific thioredoxin reductase |
| Thioredoxin reductase (TR3) | Reduction of mitochondrial thioredoxin |
| Selenophosphate synthetase 2 (SPS2) | Selenophosphate synthesis |
| 15 kDa Selenoprotein (Sep15) | Quality control of protein folding in the Endoplasmic Reticulum |
| Selenoprotein H (SelH) | Unknown |
| Selenoprotein I (SelI) | Unknown |
| Selenoprotein K (SelK) | Endoplasmic Reticulum-associated degradation |
| Selenoprotein M (SelM) | Unknown |
| Selenoprotein N (SelN) | Involved in muscle development |
| Selenoprotein O (SelO) | Unknown |
| Selenoprotein P (SelP) | Se transport |
| Methionine-R-sulfoxide reductase R (SelR) | Reduction of oxidized methionine residues |
| Selenoprotein S (SelS) | Endoplasmic Reticulum-associated degradation |
| Selenoprotein T (SelT) | Unknown |
| Selenoprotein V (SelV) | Unknown |
| Selenoprotein W (SelW) | Unknown |
Hopefully from looking at the table, you see that the glutathione peroxidase enzymes and thioredoxin reductases are antioxidant enzymes. The iodothyronine 5'-deiodinases are involved in the metabolism of thyroid hormones, which will be discussed further in the iodine section. For the vast majority of the other selenoproteins, their function is not known, so they were named selenoprotein and given a letter. As described earlier and shown below, glutathione peroxidase converts hydrogen peroxide into water.
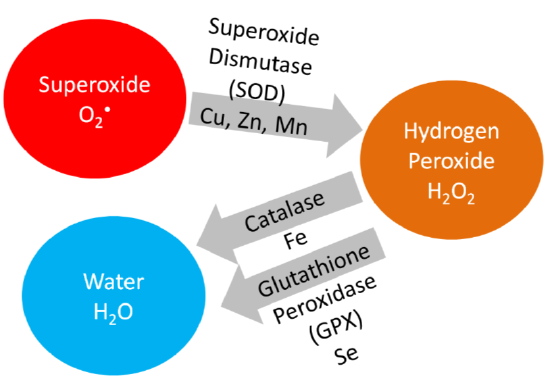
Remember that thioredoxin reductase can regenerate ascorbate from dehydroascorbate in the theorized antioxidant network (shown below).
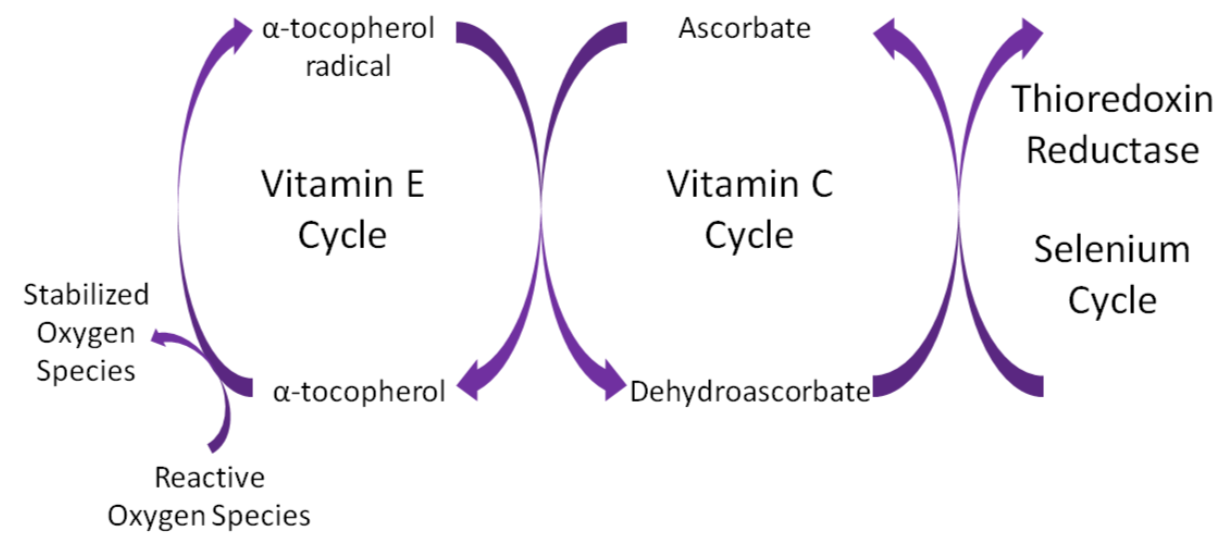
Query \(\PageIndex{4}\)
Selenium Absorption, Excretion, Toxicity & Its Questionable Deficiency
Selenium is highly absorbed. Thus, selenium levels in the body are not regulated by absorption, but rather by urinary excretion. Organic selenium forms may be absorbed slightly better than inorganic forms, as one study found that 98% of a dose of selenomethionine was absorbed, compared to 84% of selenite1.
Selenium is primarily excreted in the urine, but at high levels it can be expired, producing garlic odor breath.
Selenium toxicity can be a problem, especially for animals living in or around a body of water in an area with high soil selenium levels. This is because runoff from the soil causes selenium to collect in the water in high levels and then starts working its way up the food chain and causing problems, as shown in the following link.
Web Link
In humans, the initial symptoms are nausea, fatigue, and diarrhea. If continued, the person may develop hair and nail brittleness, rash or skin lesions, and nervous system abnormalities.
There are two questionable selenium deficiency diseases, Kashin-Beck disease and Keshan disease. Kashin-Beck disease is a disorder of the cartilage, bone, and joints that results in deformity and stunted growth. These disabling symptoms result in restricted movement and joint enlargement. The etiology of Kashin-Beck disease remains controversial as it is seen primarily in Tibet and other parts of China, Siberia, and North Korea. The main controversy with Kashin-Beck disease being linked to Selenium deficiency is that the pathogenesis is likely multifactorial in nature. Specifically, selenium deficiency alone cannot cause the occurrence of this disease. However, in combination with other health issues, it has been shown to be linked with Selenium deficiency6.
The second controversial disease is Keshan disease. This disease occurred primarily in the mountainous regions of China, causing heart lesions. Below is a topographical map of China to give you an idea of what areas are mountainous.

You can see in the link below the areas where Keshan disease occurred. For the most part, you can see that these 2 areas overlay.
Web Links
However, sodium selenate supplementation failed to totally eradicate Keshan disease like you would expect if it was caused by selenium deficiency. The incidence of Keshan disease also fluctuated seasonally and annually, which is unusual for a deficiency and more consistent of an infectious disease. Research found coxsackievirus in the heart of Keshan disease victims. They isolated this virus and used it to perform the experiment illustrated below.
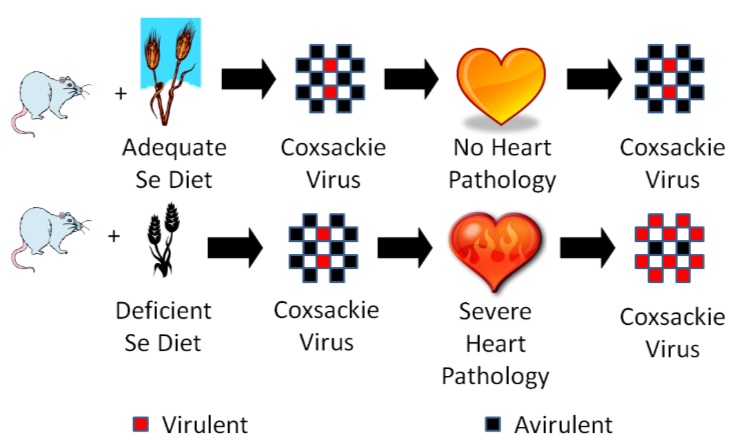
One group of mice was fed an adequate selenium diet and another group a deficient selenium diet. They were then infected with coxsackievirus that was mostly avirulent, but also contained some virulent viruses. A virulent virus is one that causes a disease, and an avirulent virus is one that does not cause a disease (some vaccines use avirulent viruses). After a period of time, they found that the selenium deficient animals developed severe heart pathology, while the selenium adequate animals did not develop heart pathology. They then isolated the virus from the hearts of the mice from both groups and found that the coxsackievirus from the deficient animals’ hearts had become mostly virulent7. They then took it one step further as shown in the figure below.
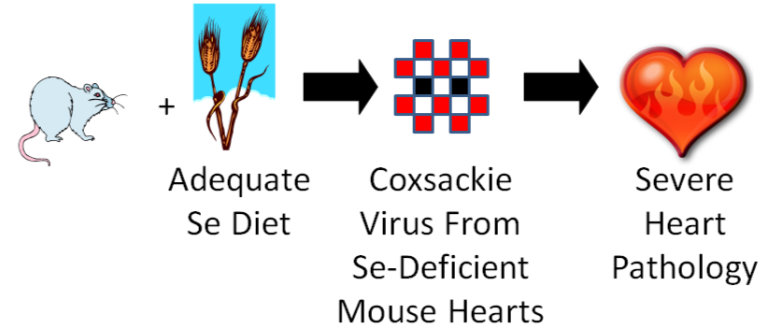
They took the isolated virus from the selenium-deficient mouse hearts and infected selenium adequate animals with it. The selenium adequate animals developed severe heart pathology like the selenium-deficient animals had previously.
What's going on? They found mutations in the virus from the selenium-deficient animals that they believe caused it to become virulent. They believe that high oxidative stress, as a result of inadequate antioxidant selenoenzymes, in these animals leads to mutations in the virus causing it to become virulent8.
Who cares? Research has found similar results with vitamin E. Researchers are also examining the effects on other viruses such as influenza (flu) and HIV. If they find a similar phenomenon occurring in other viruses, it means that you and your friend who eats a horrible diet (eats no fruit and vegetables) could be exposed to a virus. You don't know you were exposed because your immune system fights off the virus. However, your friend gets sick. He/she can serve as a host in which the virus mutates making it more virulent, which when you're exposed a second time, may make you sick.
Query \(\PageIndex{5}\)
Query \(\PageIndex{6}\)
Query \(\PageIndex{7}\)
References
- Stipanuk MH. (2006) Biochemical, physiological, & molecular aspects of human nutrition. St. Louis, MO: Saunders Elsevier.
- http://tin.er.usgs.gov/geochem/doc/a...es/se/usa.html
- Lindshield B, Ford N, Canene-Adams K, Diamond A, Wallig M, et al. (2010) Selenium, but not lycopene or vitamin E, decreases growth of transplantable dunning R3327-H rat prostate tumors. PloS One 5(4): e10423-e10423.
- Labunskyy VM, Hatfield DL, Gladyshev VN. (2014) Selenoproteins: molecular pathways and physiological roles. Physiol Rev. 94(3):739-777.
- Packer L, Weber SU, Rimbach G. (2001) Molecular aspects of alpha-tocotrienol antioxidant action and cell signalling. J Nutr 131(2): 369S-373S.
- Prabhu KS, Lei XG. (2016) Selenium, Advances in Nutrition 7(2): 415–417.
- en.Wikipedia.org/wiki/File:Ch..._35.63718N.jpg
- Beck M, Handy J, Levander O. (2004) Host nutritional status: The neglected virulence factor. Trends Microbiol 12(9): 417-423.