
7.3: Extrahepatic Macronutrient Metabolism
- Page ID
- 21005

Because the liver is so important in metabolism, the term extrahepatic has been defined to mean "located or occurring outside of the liver1". We are next going to consider extrahepatic tissue metabolism.

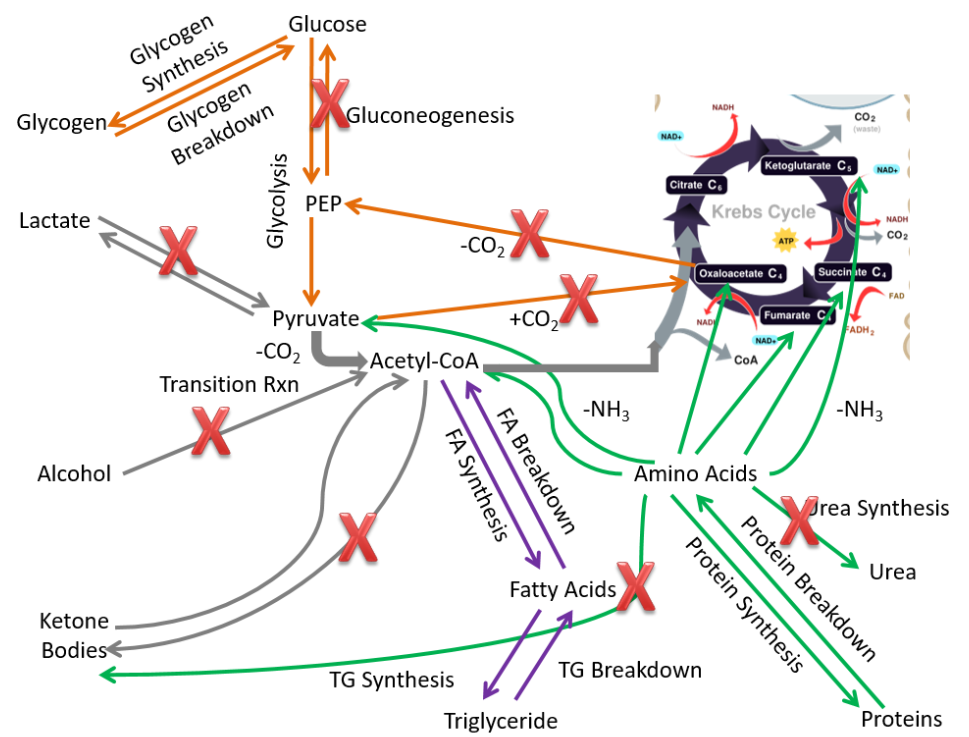
To start considering the metabolic capabilities of the extrahepatic tissues, we start by removing pathways that only or mostly occur in the liver:
- Alcohol oxidation
- Gluconeogenesis
- Ketone body synthesis
- Urea synthesis
- Lactate breakdown
- Glucose-6-phosphatase
These metabolic processes are crossed off in the figure below.
We are left with metabolic capabilities that are listed and shown below.
- Glycogen synthesis and breakdown
- Glycolysis
- Fatty acid synthesis and breakdown
- Triglyceride synthesis and breakdown
- Protein synthesis and breakdown
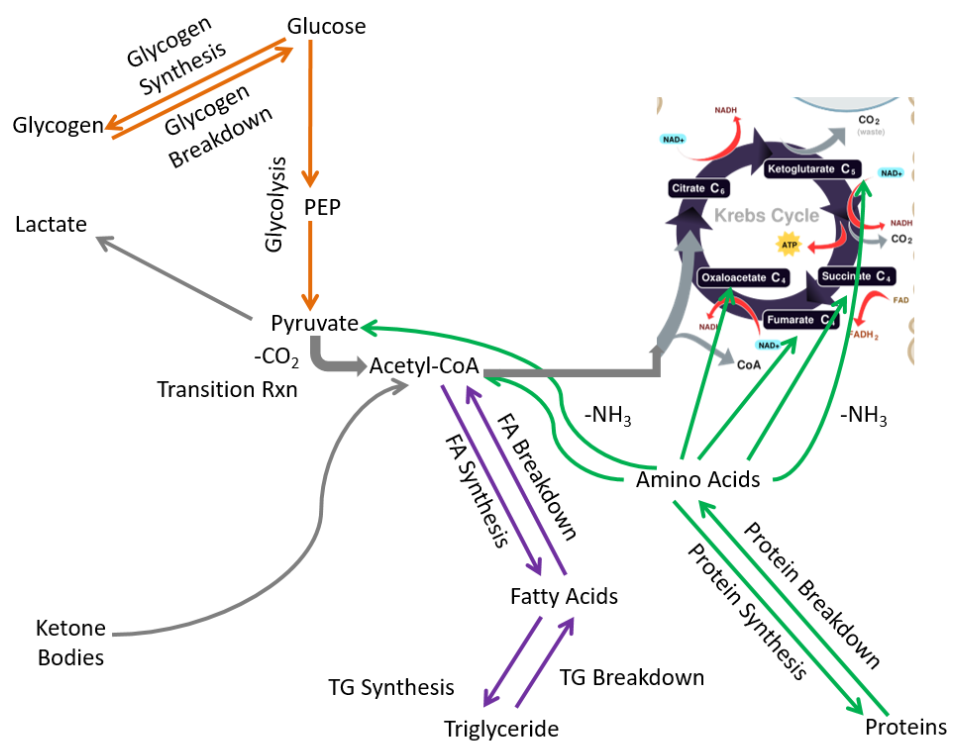
We will use this figure as the base for metabolic capabilities of the different extrahepatic tissues to compare what pathways other tissues can perform versus all the pathways performed by extrahepatic tissues.
In an effort to keep this simple, we are going to focus on four extrahepatic tissues in the following subsections:
- Muscle Macronutrient Metabolism
- Adipose Macronutrient Metabolism
- Brain Macronutrient Metabolism
- Red Blood Cell Macronutrient Metabolism
Muscle Macronutrient Metabolism
Compared to extrahepatic tissues as a whole, in the muscle the following pathways are not performed or are not important:
- Fatty acid synthesis
- Ketone body breakdown
These pathways are crossed out in the figure below.
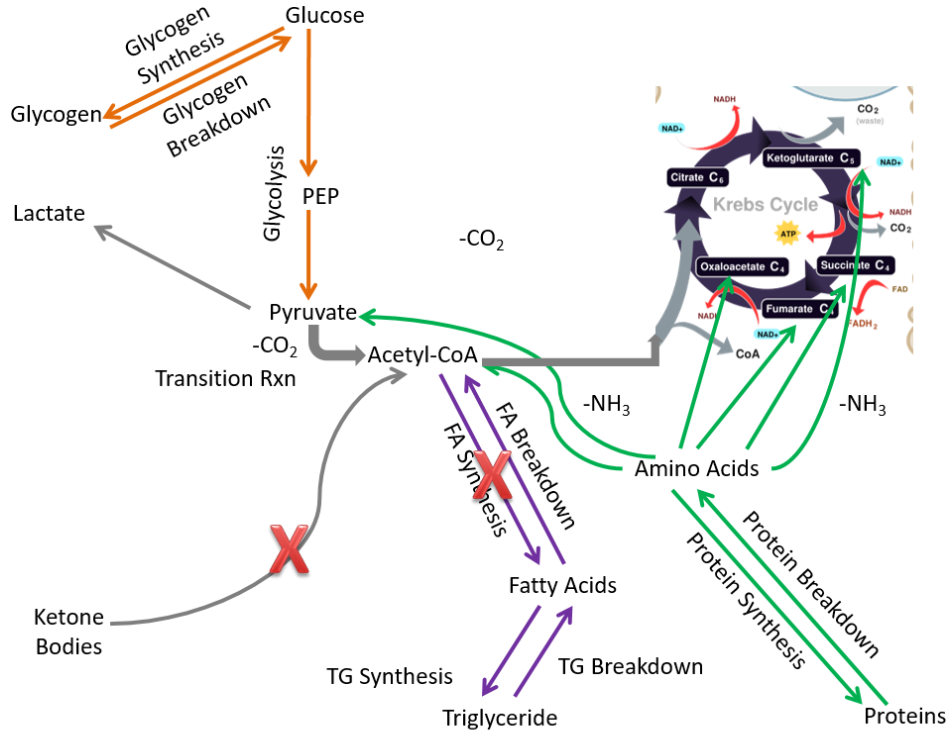
Removing those pathways, the following metabolic pathways make up the muscle metabolic capability:
- Glycogen synthesis and breakdown
- Glycolysis
- Protein synthesis and breakdown
- Triglyceride synthesis and breakdown
- Fatty acid breakdown
- Lactate synthesis
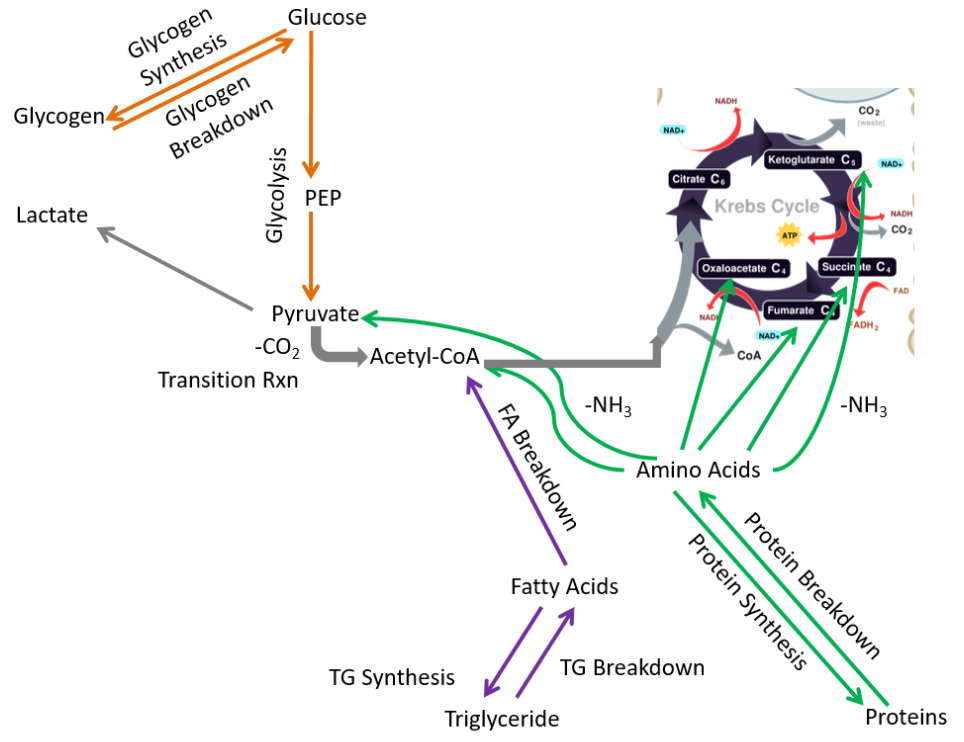
Muscle is a major extrahepatic metabolic tissue. It is the only extrahepatic tissue with significant glycogen stores. However, unlike the liver, the muscle cannot secrete glucose after it is taken up (no glucose-6-phosphatase). Thus, you can think of the muscle as being selfish with glucose. It either uses it for itself initially or stores it for its later use.
Adipose Macronutrient Metabolism
It probably does not surprise you that the major function of the adipose is to store energy as triglycerides. Compared to extrahepatic tissues as a whole, in the adipose the following pathways are not performed or are not important:
- Glycogen synthesis and breakdown
- Lactate synthesis
- Ketone body breakdown
- Fatty acid breakdown
- Protein synthesis and breakdown
- Citric acid cycle (not much since it is not an active tissue needing energy)
These pathways are crossed out in the figure below.
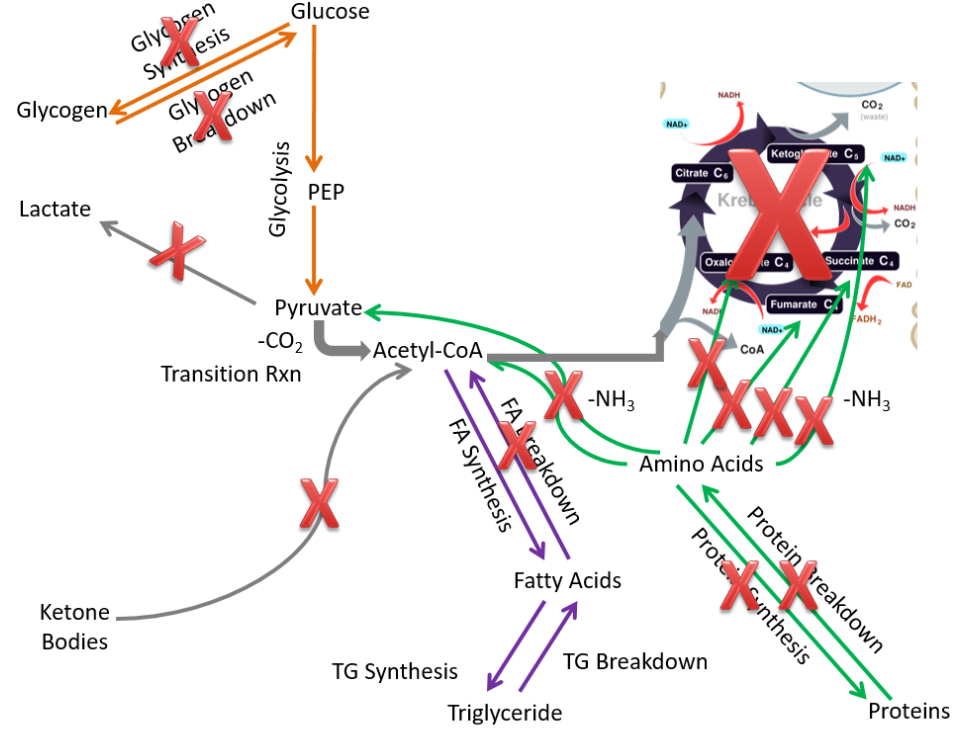
Removing those pathways, we are left with metabolic capabilities listed below and depicted in the following figure:
- Glycolysis
- Fatty acid synthesis
- Triglyceride synthesis and breakdown
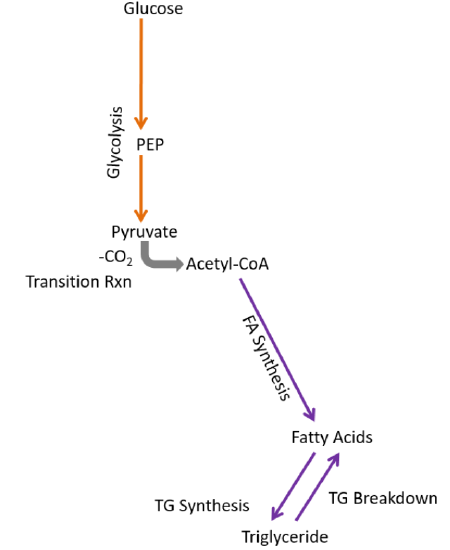
Fatty acid synthesis only occurs in the adipose and liver. In the adipose, fatty acids are synthesized and most will be esterified into triglycerides to be stored. In the liver, some fatty acids will be esterified into triglycerides to be stored, but most triglycerides will be incorporated into VLDL so that they can be used or stored by other tissues.
Brain Macronutrient Metabolism
Fatty acid breakdown does not occur to any great extent in the brain because of the low activity of an enzyme in the beta-oxidation pathway limits the pathway’s activity1. Compared to the extrahepatic tissues as a whole, in the brain the following pathways are not performed or are not important:
- Glycogen synthesis and breakdown
- Lactate synthesis
- Fatty acid synthesis and breakdown
- Triglyceride synthesis and breakdown
- Protein synthesis and breakdown
These pathways are crossed out on the figure below.
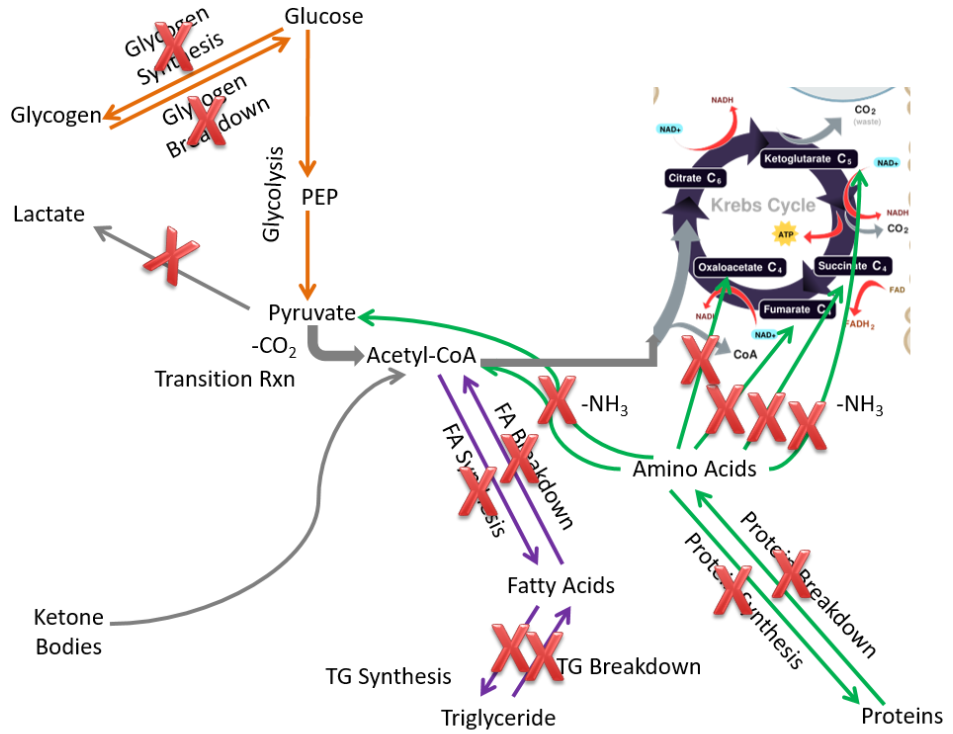
Fatty acid breakdown does not occur to any great extent in the brain because low activity of an enzyme in the beta-oxidation pathway limits the activity of this pathway4.
By removing those pathways the only pathways left are:
- Glycolysis
- Ketone body breakdown
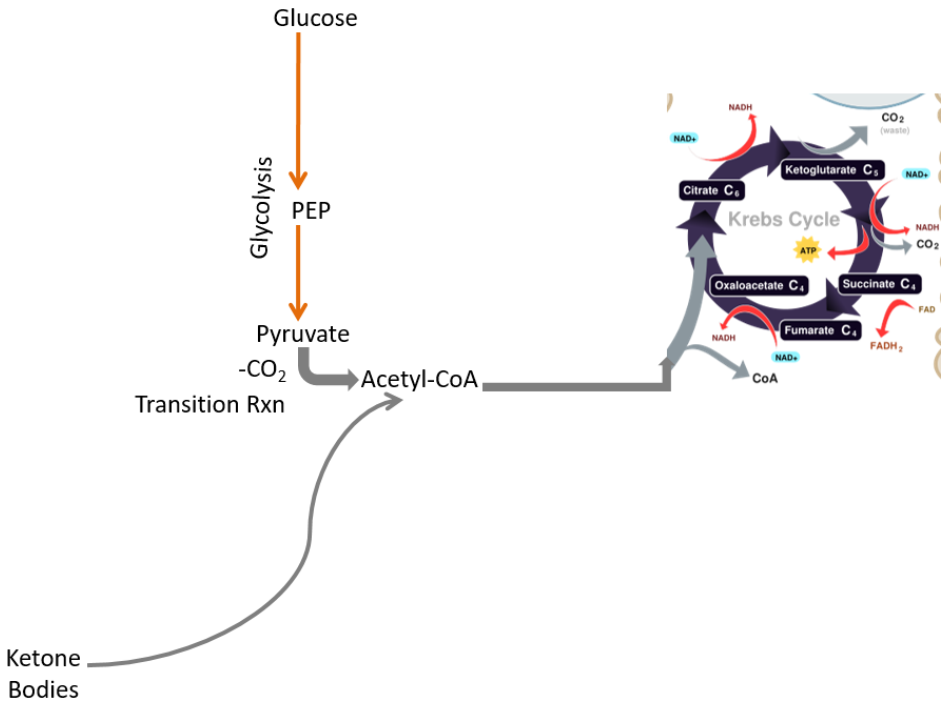
Thus, due to its limited metabolic capabilities, the brain needs to receive either glucose or ketone bodies to use as an energy source.
Red Blood Cell Macronutrient Metabolism
Red blood cells are the most limited of the extrahepatic tissues because they do not contain a nucleus or other cell organelles, most notably mitochondria.
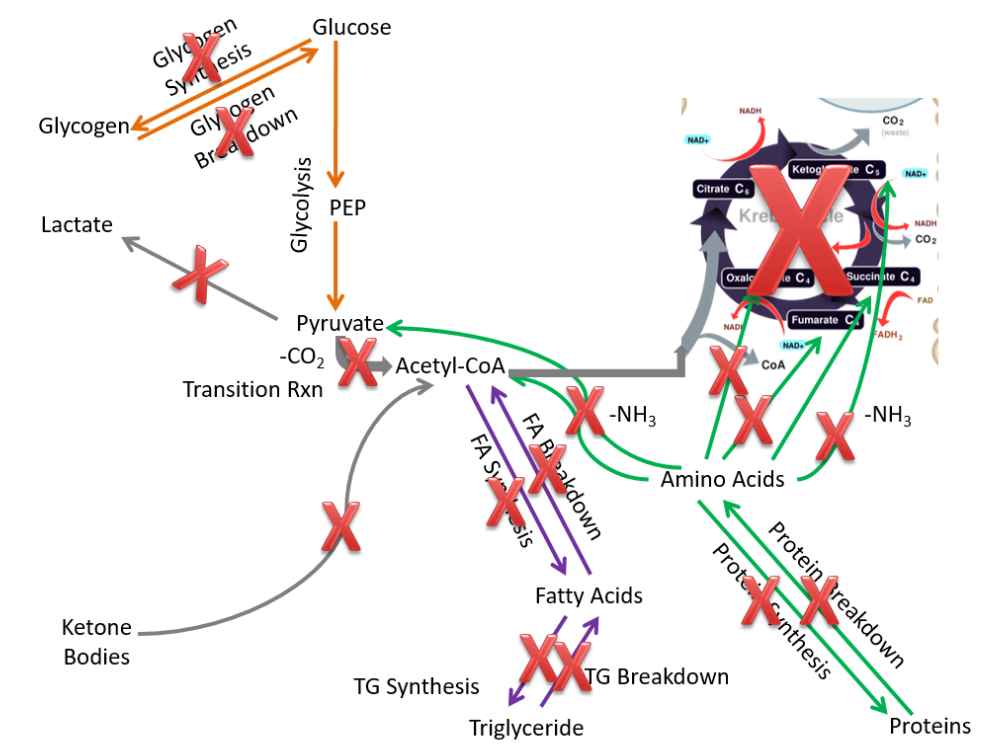
As a result, compared to the extrahepatic tissues, in red blood cells the following pathways are not performed or are not important:
- Glycogen synthesis and breakdown
- Lactate breakdown
- Fatty acid synthesis and breakdown
- Triglyceride synthesis and breakdown
- Protein synthesis and breakdown
- Ketone body breakdown
These pathways are crossed off in the figure below.
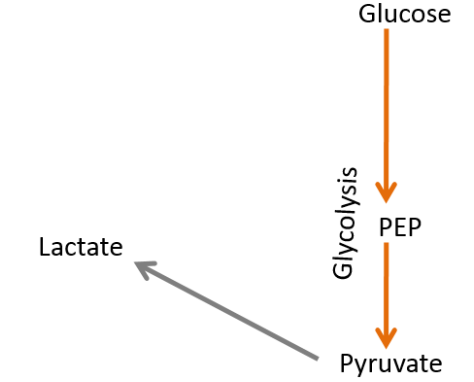
If all those pathways are removed, only glycolysis is left, where pyruvate is converted to lactate.
Thus, red blood cells are one-trick ponies, only being able to perform glycolysis and produce lactate.
References
- http://www.cancer.gov/dictionary/?CdrID=44498
- commons.wikimedia.org/wiki/File:Liver.svg
- en.Wikipedia.org/wiki/File:CellRespiration.svg
- Yang SY, He XY, Schulz H (1987) Fatty acid oxidation in rat brain is limited by the low activity of 3-ketoacyl-coenzyme A thiolase. J BIol Chem 262 (27): 13027-13032.
- en.Wikipedia.org/wiki/Mitochondrion