
1.7: Pharyngeal Arches
- Page ID
- 41543

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Overview of the pharyngeal arches
- Development of the external structures
- Fronto-nasal process
- 1st pharyngeal arch
- maxillary process
- 2nd-6th pharyngeal arch
- Pharyngeal grooves
- Pharyngeal pouches
- Development of internal structures
- Pituitary gland
- Salivary glands
- Palate
- Tongue
- Tonsils
- Clinical application of pharyngeal arch development
- Cleft lip/palate
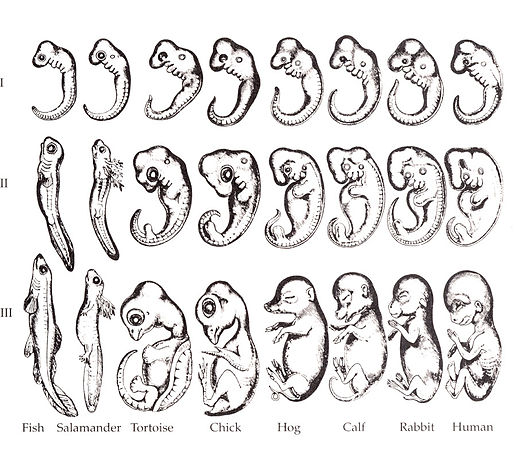
Overview of the pharyngeal arches
Ontogeny recapitulates phylogeny —Ernst Haeckel
Nothing in biology makes sense except in the light of evolution–Theodosius Dobzhansky
The human embryo is taking shape. It might seem odd that we look a bit wormy at this early stage, and next we morph into something fishy in appearance. The phrase ontogeny recapitulates phylogeny is hallowed among developmental biologists. It means, more or less, our early embryonic stages look like our evolutionary lineage. That is what the famous drawing (Fig. 7.1) shows: trace our evolutionary lineage (over millions of years), it looks like our embryonic lineage (the 9 months before birth). In this chapter, we see the human embryo grow gills arches, and re-shape those gill arches into other structures: a mandible and maxilla, ears, and salivary glands (now that we don’t have easy access to saline to lubricate every bite of food). Why not just form a mandible and ears, why go through the middle gill arch step? Why do we begin resembling worms, then fish, then only much later, little people? The better question, from the perspective of an evolutionary biologist, is why would we stop developing the way our ancestors did? Would that somehow help us avoid predation, have more offspring or have superior offspring? The answer is apparently either no, or its not worth the risks involved. Therefore, we rarely see new structures arise in evolution de novo (from the beginning), instead we observe structures changing slightly over time. This leads to different species having similarly shaped structures with potentially different functions, or homologues. A bat’s wing, a whale’s flipper, a horse’s leg and your arm have the same basic skeletal pattern: 1 bone, 2 bones, 4 bones (the thumb is different), and are referred to as homologues. Their size, shape and purpose are different, but you don’t need an entirely new design, it is more efficient to morph a leg into a flipper. Some homologues have very different functions, such as human lungs and fish swim bladders. Conversely, the wings of a fly, chicken and bat, despite their similar function, are not homologous, nor is the panda’s (6th) thumb a homologue of the human pollex. This all becomes apparent when you study the lineage of different species and compare it to their development.
Homologues also exist within a single species. That is why your arm and leg share the same skeletal pattern, they too are homologues. Sharing instructions for different structures in the human body cuts down on the amount of DNA that needs to be built and copied with each cell division. Small changes in DNA can lead to big changes in morphology. Therefore, it is both faster and takes less space to tweak a design that works than create a whole new design de-novo. Human development is rife with examples of recapitulation, where the same basic process is re-used with small changes.
Keep in mind we did not evolve from modern-day worms or fish, but the most recent relative a trout and ourselves share resembled the fish more than us, and the most recent relative a trout and worm share resembled the worm more than the trout. Now reverse that: our embryonic stages initially look strikingly like some sort of parasitic worm, then more fishy, then kind of lizardy, and finally mammally. Another way to say that is ontogeny recapitulates phylogeny.

If the link between of the evolution and development seems complicated, it is. The purpose of this overview section is to conceptually prepare you for a complicated series of steps that go through unexpected transitions. Evolution explains why the transitions happen, your job is to learn what those transitions are.
Before we start, however, we would like to point out one more level of complexity. You need to differentiate between structures that have similar names, including the pre-maxillary segment and inter-maxillary segment, the mandibular arch and maxillary process, the maxillary process and the inter-maxillary segment, the facial prominences and pharyngeal arches, pharyngeal grooves and pharyngeal pouches, and more. Furthermore, most of these structures have multiple names, such as the pharyngeal arches, gill arches and branchial arches, which can make comparing this text to others tricky.
If at any time you feel like you need more detail or a different description, let us remind you of this excellent web resource:
- The Embryology Education page
- by Dr. Mark Hill at the University of New South Wales, Sydney, Australia
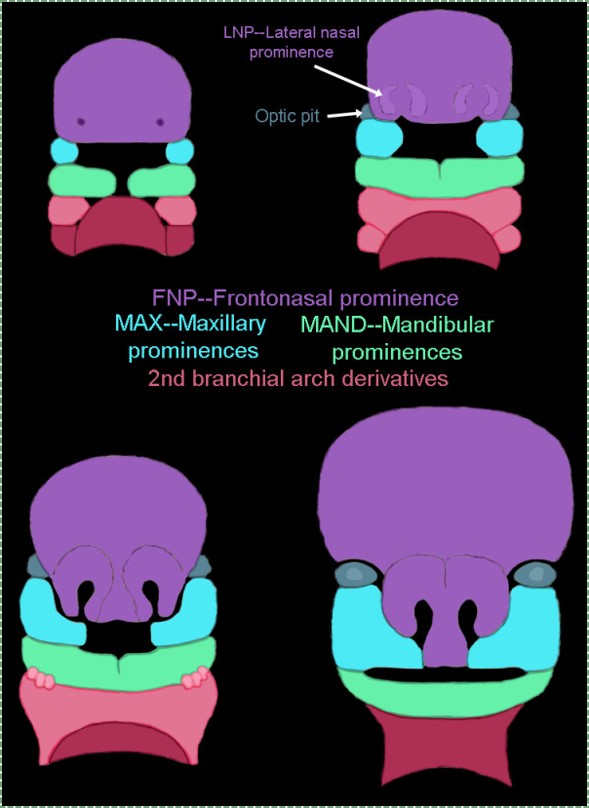
Let us return our focus to human embryos. At 4 weeks, the face begins to develop. It is composed of several parts (or prominences), listed in Table 7.1. To use the word prominence means these are not necessarily the same type of thing, just things we can see. Some prominences are homologues of gill arches (gill arches are in Fig. 7.2, their human homologues are in Fig. 7.3 upper left). They are quickly remodeled (Fig 7.3, lower right). These pharyngeal arches (or branchial arches, although the name branchial arches should be limited to vertebrates with gills), are paired segmental bulges on the lateral borders of the primitive pharynx. Each pair of arches grows medially across the face (the ventral side) and fuses with its partner. One pair of processes (not arches) tries to fuse, but run into a bump called the fronto-nasal process, and fuse with it instead. If we were lampreys, the arches wouldn’t fuse at all, and remain 6 gill arches. If we were bony fish, we would remodel 3 arches and have a mandible, inner ear structures and 3 gill arches. As humans, the pharyngeal arches are fated to become a number of cool things, including teeth, but not gill supports.
| The facial prominences |
|---|
| Mandibular (1st) arch |
| Maxillary process |
| Fronto-nasal process |
| Hyoid (2nd) arch |
Table 7.1: The facial prominences include two pharyngeal arches.
Development of the face (external)

Formation of the pharyngeal arches
At the 4th week, the primitive gut has not fused with ectoderm, so no oral or nasal cavities exist. Where the mouth will form is visible as a depression in the ectoderm known as the stomodeum (the primitive mouth). A thin membrane, the oro-pharyngeal membrane (or bucco-pharyngeal membrane), separates the stomodeum from the end of the primitive foregut that becomes the pharynx. The primitive foregut is lined by endoderm.
The pharyngeal arches form under the instruction of homeobox genes. Activation of homeobox genes (in an anterior-to-posterior pattern) induces a program of other genes to be transcribed, including morphogens which induce growth of all 3 germ layers. This causes outward budding, creating pharyngeal grooves between each pair of pharyngeal arches on the outside of the embryo. On the inside of the primitive pharynx, opposite of the pharyngeal arches, localized growth forms invaginations known as the pharyngeal pouches. If you remember GAP, this mnemonic may help you to remember the names of these structures from external to internal: Groove, Arch, Pouch. Or, if you prefer the name pharyngeal cleft rather than groove, the mnemonic becomes CAP. These structures appear one pair at a time, from anterior to posterior, and their fate is listed in Table 7.2. Because the arches are quickly remodeled into other structures, we say that they are transient structures, but their brief existence explains how and why the adult structures in Table 7.2 have the morphology that they do.
| Arch # | Name | Ectoderm and neuro-ectoderm fate | (associated groove) | Mesoderm and neuro-mesenchyme fate | Endoderm (pouch) fate |
|---|---|---|---|---|---|
| 1st | Mandibular arch | Maxillary process –> upper lip epidermis | n/a
(this is not an arch) |
Dermis, maxilla, zygomatic, palatine, vomer | n/a
(this is not an arch) |
| Lower lip epidermis, Trigeminal nerve (CNV). | External acoustic meatus | Dermis, mandible, malleus, incus | Eustachian tube | ||
| 2nd | Hyoid arch | Epidermis, Facial nerve (CNVII) | disappears | Dermis, most of the hyoid bone, stapes | Palatine tonsils |
| 3rd | Epidermis, Glossopharyngeal nerve (CNIX) | Dermis, the rest of the hyoid bone | Thymus, Parathyroid glands | ||
| 4th | Epidermis, Vagus nerve (CNX) | Dermis, Thyroid cartilage, epiglottis | Parathyroid, Thyroid glands | ||
| 5th | disappears | ||||
| 6th | Epidermis, Vagus nerve (CNX) | disappears | Dermis, the other laryngeal cartilages | Larynx tissues | |
Table 7.2: The pharyngeal arches and their fate, separated by the germ layers.
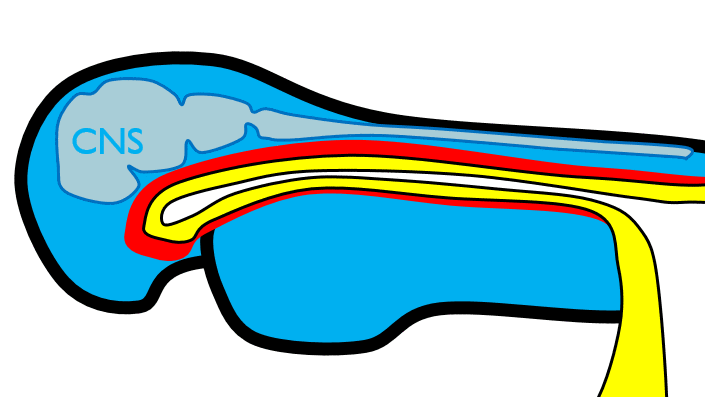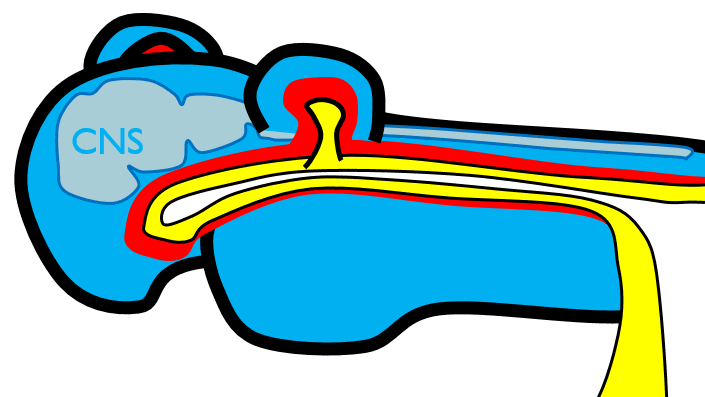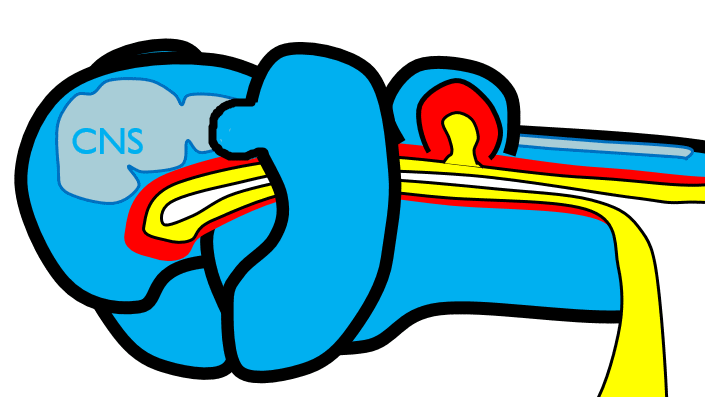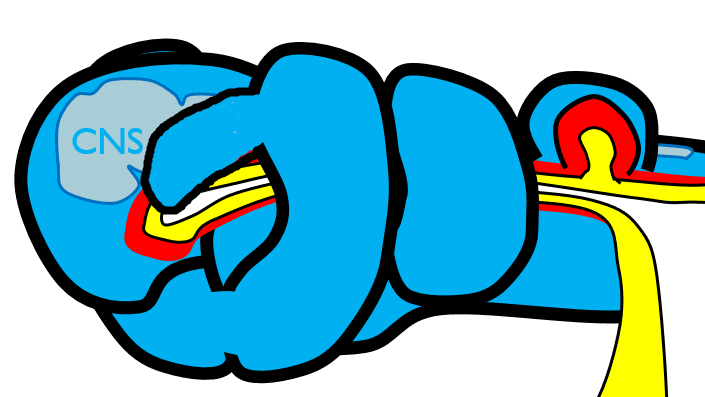
The 1st pharyngeal arch
Next, neural crest cells guide modification of the pharyngeal arches. Neural crest cells first migrate from the head into pharyngeal arch mesoderm, undergo an epithelial-to-mesenchymal transition, and differentiate into special mesenchymal stem cells called neuro-mesenchymal stem cells. The neuro-mesenchymal stem cells release different morphogens which induce localized growth within the mandibular arch (the first pharyngeal arch), forming the maxillary process. The rest of the mandibular arch grows medially, as does the maxillary process, to form the upper and lower jaw. They grow above and below the stomodeum, which at this time is still not open to the primitive pharynx.
Tissue also grows on either side of the nasal placodes. Two growths in the middle, the medial nasal processes, fuse to form the inter-maxillary segment (or globular process). The growths on the sides form lateral nasal processes, which become the alae of the nose.

Fusion of the 1st pharyngeal arch
The mandibular arch forms on the lateral edges of the embryo during the 4th week of development and grows medially. The two halves fuse by the end of the 4th week of development, creating a single structure that becomes the mandible and nearby tissues. For the two halves of the mandibular arch to grow medially, mesenchyme is removed. This requires induction of the gene for the enzyme hyaluronase, which digests Hyaluronic Acid found in ground substance. This allows epithelial cells to fuse with epithelial cells from the other half of the arch. Fusion requires matching cell-adhesion molecules and desmosomes. The mesoderm of one arch also fuses with mesoderm of its partner, which requires matching the correct integrin to fibronectin.
Later, neuro-mesenchyme of the first pharyngeal arch differentiate into a cartilaginous structure known as Meckel’s cartilage. Parts of Meckel’s cartilage undergo endochondral ossification to become part of the mandible and middle ear bones, the rest undergoes apoptosis. The mandible forms from 4 separate parts beginning in the 10th week of development, which is more complicated than most bones. The body develops by intra-membranous ossification formed from the dense connective tissue derived from mesoderm. The rami (including coronoid and condylar processes) grow by endochondral ossification of Meckel’s cartilage, derived from neuro-mesenchyme. This creates a cartilage-containing synovial joint between the ramus and temporal bones (the temporo-mandibular joint), and a fibrous joint between the two halves of the body which fully ossifies.
Fusion of the maxillary processes and inter-maxillary segment
The pair of maxillary processes grow medially in the 4th week, but run into the inter-maxillary segment and fuse with it by the 10th week of development. The upper lip, therefore, is formed of three parts (the left and right maxillary processes, and the inter-maxillary segment) while the lower lip just two (the left and right mandibular arches). The philtrum is the middle section derived from the medial nasal process. It does not serve a function in humans, it just happens to be there because of how we develop (kind of like the choice of wood usd in the record player shelf). Many anatomy textbooks have a deep love of describing the functions of organs based on their adult form. Some that have tried to describe the function of the human philtrum based on its shape and location. They should learn to love development (ontogeny) and evolution (phylogeny)– anatomy makes more sense that way.

Fate of the pharyngeal grooves and pharyngeal pouches
If you were a fish, most of the pharyngeal grooves would develop into gills. As creatures of the land, grooves are either filled in or develop into other useful structures. Between the mandibular and hyoid arches, the first pharyngeal groove invaginates further and forms a tube that becomes the external acoustic meatus. It is lined by ectoderm. The other pharyngeal grooves disappear. One the opposite side, within the primitive pharynx, the pharyngeal pouches invaginate and grow towards the grooves. The first pharyngeal pouch elongates into a tube that is fated to become the Eustachian tube, connecting the pharynx to the middle ear (the internal acoustic meatus forms when bone tissue grows around cranial nerve VIII, connecting the inner ear to the brain). The Eustachian tube is lined by endoderm. The mesoderm and neuro-mesenchyme between these two tubes forms middle ear structures, including the malleus, incus and stapes bones.
The other pharyngeal pouches invaginate and form tonsillar and glandular tissue. Tonsils are lined by endoderm-derived epithelium, but the white blood cells of the germinal centers migrate there from bone marrow stem cells (which, like most connective tissues, are dervied from mesoderm).

Development of the salivary glands
The salivary glands develop in a process that begins similarly to the formation of the teeth and neural tube. Placodes form on the ectoderm starting between weeks 4 through 12, under the control of morphogens including members of the FGF family. Salivary gland placodes grow and invaginate from there. Eventually, stem cells differentiate into a number of different epithelial cell types. The ducts are predominantly simple cuboidal epithelia. These epithelial cells have different functions based on their distal-to-proximal location along the duct. The differentiation of salivary gland cells along a proximal-to-distal axis is guided by planar cell polarity morphogens, including members of the Wnt family. Some cells in the acini differentiate into myo-epithelial cells. Myo-epithelial cells are ectodermal by lineage, and therefore epithelial, despite looking and acting like smooth muscle cells. Myo-epithelial cells express genes mostly used only by muscle cells. This involves removing histones and DNA methylation from genes shut down in cells as they initially adopted an epithelial fate.
Development of the palate and other internal structures
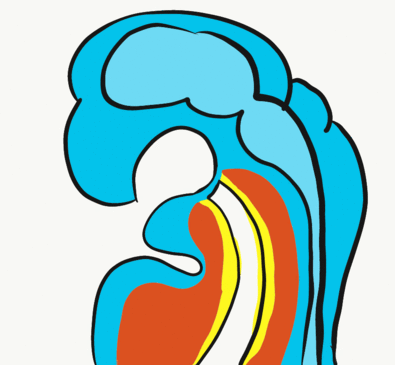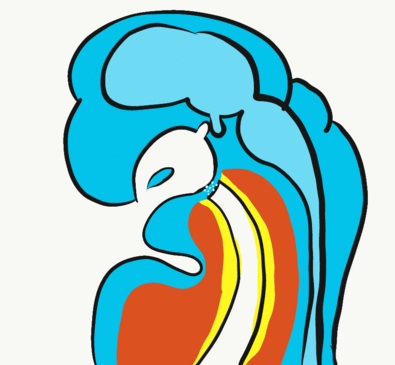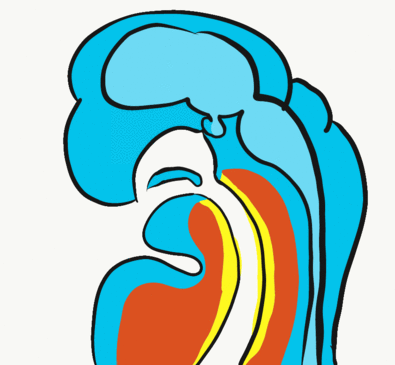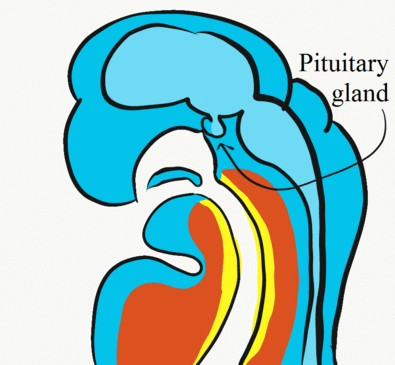
Formation of the pituitary and mouth
Inside the stomodeum, a single invagination of ectoderm forms, along the medial portion of the roof (so far, all of the processes and pouches have been left/right pairs). This invagination is named Rathke’s pouch. It grows and meets a downward budding of neuro-ectoderm. These two fuse to form the pituitary gland. The ectoderm forms the glandular half (adenohypophysis), and the neuro-ectoderm forming the infundibulum and neural half (neurohypophysis) of the pituitary gland. Rathke’s pouch fills in as the two halves of the pituitary fuse, but it is possible a small depression will remain.
At the same time, the oro-pharyngeal membrane undergoes apoptosis. This connects the primitive pharynx and stomodeum, forming the primitive oro-nasal cavity. Finally, the mouth and anus are connected! The oral mucosa therefore develops from ectoderm of the stomodeum, while the pharynx and ventral surface of the tongue are derived from endoderm.

Formation of the palate
Shortly after the lips begin forming, the palate begins to form as well, dividing the primitive oro-nasal cavity into a more mature oral cavity and nasal cavities. The palate has 3 parts that fuse with each other, and with the nasal septum. The primary palate grows from the inter-maxillary segment, and two palatal shelves grow from the maxillary processes.
| Structure | Lineage | Forms during | Fuses with |
|---|---|---|---|
| Primary palate
(pre-maxillary segment) |
inter-maxillary segment (globular process) | 6th week | Secondary palate: 9th week |
| Secondary palate
(Palatal shelves) |
Maxillary process | 7th week | The other palatal shelf: 9th week |
| Primary palate: 9th week | |||
| Nasal septum: 12th week |
Table 7.3: Summary of the development of the palate.

The first part of the palate to form is the primary palate, which develops from the inter-maxillary segment. When it forms initially, it partially divides the future oral and nasal cavities. Next, two palatal shelves grow off of the maxillary processes (Fig. 7.11). The palatal shelves first grow inferiorly, then change direction and grow medially. At this time, the developing tongue must move out of the way. This allows the palatal shelves to meet and fuse with the primary palate, as well as each other. The fusion happens in an anterior-to-posterior direction. All of this growth is directed by morphogens, including Fibroblast Growth Factors, Bone Morphogenetic Proteins, and Sonic Hedgehogs.
Maxillary incisors develop from the primary palate, while maxillary canines, pre-molars and molars develop from the secondary palate. At the site where the primary palate and the two palatal shelves fuse, a small hole remains, the incisive foramen, which houses the nasopalatine artery and vein as well as a branch of the trigeminal nerve. The oral mucosa above this foramen has a bump named the incisive papilla, which shares more in common with olfactory epithelium than it does mucosal epithelium (it is the homologue of the vomeronasal organ found in many vertebrates). Where the two palatal shelves fuse leaves a ridge named the median palatine suture. This creates a visible ridge on the overlying oral mucosa called the (median) palatine raphe.
Keep in mind that we are referring to the entire palate. Much later, anterior portions of the palate mesoderm undergo endochondral ossification and form the palatine bones and the palatine processes of the maxilla (the hard palate). The rest of palatal mesoderm differentiates into muscle tissue, forming the soft palate. Time out for spelling: this is the palate, not an artist’s palette of colors, nor a pallet used in shipping, not even a plate on which we place a tasty dinner. Therefore, foodstuffs shipped on a pallet, cooked by a chef with a harmonious palette, served to us on a plate, will be enjoyed for their flavor when they hit our palate because we have a refined palate (an appreciation for flavor). Got it? English is fun.
The nasal septum grows inferiorly at this time. It fuses with the completed palate around the 12th week of development. This creates paired nasal cavities. Initially, mesoderm differentiates into the ethmovomerine cartilage, and then partially undergoes endochondral ossification to generate a bony portion (portions of the ethmoid and vomer bones) as well as cartilaginous portion. Ossification begins from a lateral pair of ossification centers, therefore the early septal bones develop as 2 layers (lamella) which fuse to form a single bony septum.

Development of the tongue
The tongue is a hybrid structure, it forms from multiple parts making its development complicated. Tongue development begins during the 4th week, after the pharyngeal arches fuse. The tongue develops from first 4 pharyngeal arches (skipping the 2nd, except for a connection to CNVII). Formation of the tongue involves proliferation, followed by fusion, followed by apoptosis of some tissue to give the tongue mobility.

At the 4th week, the pharyngeal arches fuse along the floor of the future oral cavity. A single triangular-shaped tuberculumimpar proliferates off the 1st arch, followed by two laterallingual swellings Because they come from the first pharyngeal arch, their inner lining is not endoderm like the other arches, but ectoderm from the oro-pharyngeal membrane. As these swellings grow, the 3rd and 4th arch develop a swelling named the copula, which grows over the 2nd arch. The oral mucosa of the anterior 2/3 of the tongue is derived from the ectoderm of the 1st pharyngeal arch, the posterior 1/3 from the endoderm of the 3rd pharyngeal arch, and the base of the tongue from endoderm of the 4th pharyngeal arch. Fusion of these structures occurs during the 8th week. The median lingual sulcus forms, where the left and right lateral lingual swellings fuse. The sulcus terminalis forms where the 1st and 3rd pharyngeal arches fuse. This border between the anterior and posterior portion of the tongue is pronounced due to the different lineages of the surface mucosa.

Apoptosis of tongue tissue on the ventral side leaves the tongue attached at the base, and freer to move around. Apoptosis does not remove all the tissue on the anterior portion, a small amount of mucous membrane remains, named the lingual frenulum. An invagination forms posterior to the sulcus terminalis and grows deeper, forming the thyroid gland. This process is similar to the way the anterior pituitary forms. It leaves behind a small depression named foramen cecum, which is a confusing name because foramen means hole, but this foramen fills in most of the way, making it more of a pouch. Similar to Rathke’s pouch, it serves no purpose in humans, it’s a remnant of the way epithelial tissue proliferates.
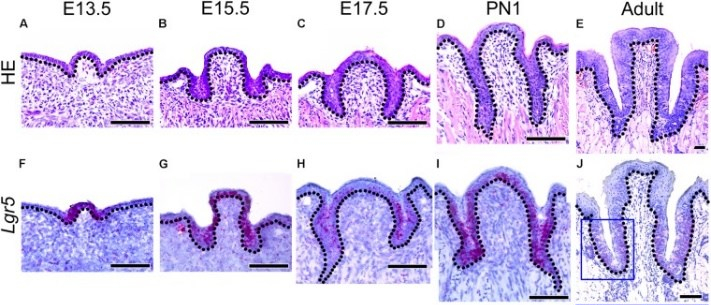
Development of the lingual papillae
The lining of the oral cavity is derived from ectoderm, the pharynx from endoderm, and the lining of the tongue is a mixture of the two. The filiform and fungiform papillae develop from invaginations of the ectoderm, while the foliate and circumvallate from invaginations of endoderm. The growth and differentiation of the papillae is guided by morphogens secreted by underlying neuro-mesenchyme, including members of the FGF and Wnt families. Keratinocytes develop from an ectodermal precursor, while taste buds (including those in the soft palate and pharynx) are induced to develop from ectodermal or endoderm precursors starting the 8th week of development. Older evidence suggests taste bud differentiation depended on neural connections, but newer evidence suggests taste buds develop in response to the Sonic Hedgehog morphogen. By adulthood, both keratinocytes and taste bud cells continue to develop from a shared epithelial stem cell, and both are replenished throughout life. Whether the lineage of this stem cell is endodermal, ectodermal or both is unclear.
The connective tissue (lamina propria, sub-mucosa, and vasculature) of the tongue is derived from neuro-mesenchyme. The skeletal muscle tissue is derived from somatic mesoderm, guided by morphogens secreted from the neuro-mesenchymal stem cells of the neuro-mesenchyme.
Clinical applications of pharyngeal arch development

Lip pits
Incomplete fusion of the pharyngeal arches leads to a number of conditions, some more severe than others. Two benign conditions include a lower labial pit, which forms when the two mandibular arches fail to fuse completely.

Commissural lip pits may form between the maxillary processes and mandibular arches. These are examples of cosmetic variations rather than congenital malformations.

Cleft lip
Incomplete fusion of either maxillary process with the inter-maxillary segment leads to the formation of a cleft lip. This can occur either on the left, right (unilateral) or both (bilateral) borders of the philtrum, although a left unilateral cleft lip is the most common. Cleft lips are more common and more severe in male children. A cleft lip may be accompanied by a cleft palate.
Cleft lip and palate occur in about 1 in 1000 births, making them a relatively common congenital malformation. Risk factors include older mothers, mothers who smoke during pregnancy or take certain medications (e.g. some anti-consultants). There are many genetic risk factors for cleft lip and palate, some examples are listed in Table 7.4.
| Gene name | Class of gene | Function |
|---|---|---|
| IRF6 | Transcription factor | Induced during development of mesoderm. |
| MSX1 | Homeobox transcription factor | Limb patterning |
| BMP4 | Morphogen | Induction and patterning of bone tissue, teeth and limbs |
| FGF10 | Morphogen | Induction and patterning of connective tissue |
| Hyal2 | Digestive enzyme | Digests hyaluronic acid prior to fusion of the lip or palate |
| p63 | Transcription factor | Controls desmosome protein expression during fusion of the lip or palate |
| Epithelial Cadherin 1 | Cell adhesion molecule | Allows epithelial cells to connect during fusion of the lip or palate |
Table 7.4: A partial list of genes that, when mutated, contribute to the formation of cleft lip/palate.
A cleft lip can cause difficulty with nursing, as it hinders the formation of a good seal around a nipple. With proper instruction, babies with cleft lip can be breast-fed or bottle-fed using a regular bottle. A cleft lip may cause problems with learning speech. Learning to speak requires sound mimicry, and because a cleft lip alters vocal sounds, it interferes with successful mimicry. Speech and hearing therapy help alleviate these problems. An increased risk of oro-nasal infections is also a concern. The preferred treatment for cleft lip is to seal the gap with surgery at 10 weeks of age. Surgery can leave behind a scar, but otherwise is highly successful.

Cleft palate
Incomplete fusion of the primary palate and/or the palatal shelves leads to a cleft palate. A cleft palate may or may not be accompanied by a cleft lip. Cleft palate is more common in females.
Cleft palate causes difficulty with nursing, because a child cannot create suction with an opening from the oral cavity into the nasal cavity. There are a number of specialty bottles that help babies with cleft palate bottle feed. Similar to cleft lip, a cleft palate can lead to difficulty learning speech. Disruption of palate formation may lead to shape changes in the Eustachian tube, which in turn alters the ability to regulate middle ear pressure and leads to an increased risk of hearing loss.

The preferred treatments for cleft palate include Naso-Alveolar Molding (NAM), followed by several surgeries. NAM involves screwing or taping an appliance to the maxilla at around 10 months of age to slowly pull in an anterio-medial direction, towards the inter-maxillary segment. Using such an appliance reduces the amount of surgery required to correct the cleft, relying more on guided growth of the child’s tissues. The device is adjusted by an orthodontist every two weeks for about a year. Multiple surgeries follow in the treatment of cleft palate– it is a complicated region, made all the more complex by the fact the child is growing fast. It is not ideal to wait for a child to stop growing for the same reason early intervention for cleft lip was important: orofacial clefts hinder speech development.
Orofacial clefts are categorized first as being a cleft lip (CL), a cleft palate (CP), or a cleft lip and palate (CL/P). A unilateralcleft lip or palate affects just the left or right side, while a bilateralcleft affects both sides. An incompletecleft palate involves incomplete fusion between the primary palate and a palatal shelf, while a completecleft also involves incomplete fusion between the two palatal shelves.
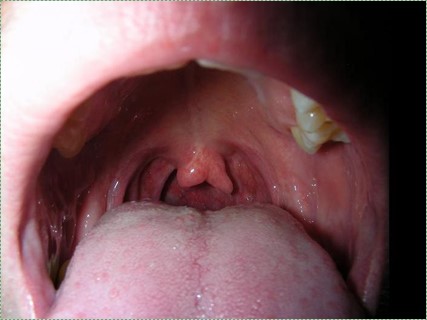
Cleft uvula
A cleft uvula is the least complicated form of cleft palate, and should be considered a cosmetic variation rather than a congenital malformation. A cleft uvula still closes off the nasopharynx during swallowing.
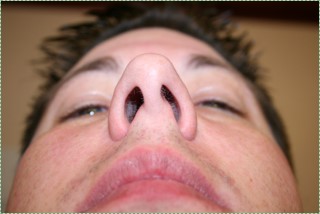
Deviated nasal septum
If the nasal septum grows at an angle as it is developing, it leads to a deviated septum. In fact, it is rare for the septum to develop in a symmetrical fashion. 80% of people have some nasal septum deviation, usually without symptoms. Complications can arise because of nasal cavity physiology. The paired nasal cavities contain erectile tissue (areolar connective tissue) below the mucosa. This tissue undergoes a nasal cycle, alternating between one side swelling shut and the other remaining open for breathing. This prevents the nasal cavities from drying out from constant use. But for someone with a significant nasal septum deviation, it leads to difficulty breathing when the larger cavity swells shut. A relatively simple surgery called septoplasty can be done to increase the size of the smaller nasal cavity. Septoplasty is not the same as the plastic surgery procedure rhinoplasty, where the shape of the nose is altered.

Palatal torus
Excessive growth of the palatal shelves can create a palatal torus (or torus palatinus), another example of a cosmetic variation. A palatal torus requires no treatment, as it usually does not cause any health-related issues, aside from complicating the fit to dentures. About 20-30% of the population has some degree of palatal torus. A mandibular torus, on the other hand, is not congenital. One can develop later in life, often as a response of bone tissue to bruxism.
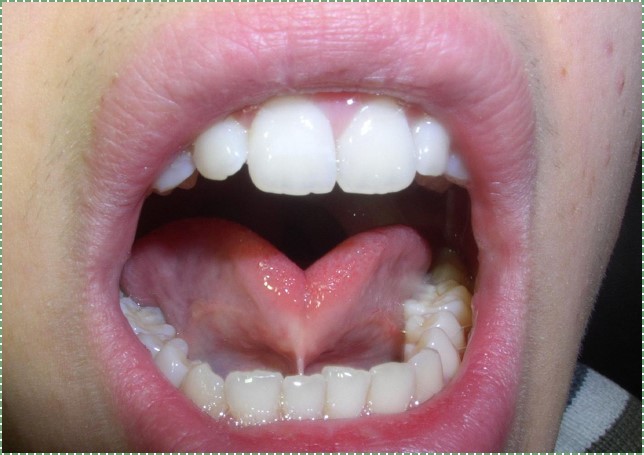
Ankyloglossia
Ankyloglossia (tongue tie) is the persistence of tissue anchoring the tongue to the floor of the mouth. Most of the ventral side of the 1st pharyngeal arch should undergo apoptosis, leaving behind a small lingual frenulum. However, with inadequate apoptosis, a pronounced lingual frenulum results, limiting the mobility of the tongue. This causes problems with breastfeeding and learning speech, but it is correctable with a minor surgery (a lingual frenectomy). Ankyloglossia is a common congenital malformation, although there is significant amount of disagreement as to how prevalent it is, with estimates ranging widely, from 1% to 25% of births (that is potentially a lot of surgical procedures). Similarly, there is disagreement as to how necessary surgical intervention is, and this disagreement has been ongoing for well over 75 years.

Pharyngeal cleft cyst
Pharyngeal cleft cysts form when incomplete fusion of neighboring pharyngeal arches leaves the remnant of a pharyngeal groove. These usually form a painless mass in the neck, until an infection occurs. They may be left untreated, or may be removed by surgery. This involves removing the extraneous ectodermal (epithelial) tissue trapped deeper in the neck. Whether surgery is or isn’t performed may depend on how close the cyst is to the carotid artery, internal jugular vein or facial nerve.
Rathke’s cleft cyst
Similar to pharyngeal cleft cysts, a cyst may form from incomplete obliteration of Rathke’s pouch during formation of the pituitary gland. This leads to a mucus-filled cyst near the anterior pituitary. Due to its location, it may put pressure on the optic chiasm, leading to visual disturbances, otherwise it is asymptomatic. Drainage is the preferred treatment over removal, owing to how close it is to the pituitary gland.
Chapter 6 * Chapter 8