
3.1: Synapse Structure
- Page ID
- 69847
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Glossary Terms
- Key Takeaways
- Test Yourself
For the nervous system to function, neurons must be able to communicate with each other and they do this through structures called synapses. At the synapse, the terminal of a presynaptic cell comes into close contact with the cell membrane of a postsynaptic neuron.

Synapse Types
There are two types of synapses: electrical and chemical.
Electrical
Electrical synapses are a direct connection between two neurons. Imagine two neurons that are connected by an electrical synapse. First of all, both of them are complete cells on their own. Each one contains a complete plasma membrane surrounding the neuron, a nucleus, and all the individual organelles needed to carry out that cell’s basic life processes.
Cell membrane proteins called connexons form gap junctions between the neurons. The gap junctions form pores that allow ions to flow between neurons, so, as an action potential propagates in the presynaptic neuron, the influx of sodium can move directly into the postsynaptic neuron and depolarize the cell. The response in the postsynaptic cell is almost immediate, with little-to-no delay between signaling in the pre- and postsynaptic neurons.
Animation 11.1. Membrane-bound proteins called connexons form gap junctions between presynaptic and postsynaptic neurons. This allows for direct exchange of ions between neurons. An action potential in the presynaptic neuron will cause an immediate depolarization of the postsynaptic membrane because the sodium ions will cross the membrane through the gap junctions. ‘Electrical Synapse – Ion Flow’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC BY-NC-SA) 4.0 International License. View static image of animation.
Since the gap junctions allow diffusion of ions without any obstruction, the signal can flow bidirectionally through an electrical synapse. The electrochemical gradients will drive direction of ion flow.
This means that a signal does not always move sequentially from the presynaptic cell to the postsynaptic cell. Rather, ions and signaling molecules are free to move through the connexons in either direction. Also, each cell within an electrically-coupled network can receive inputs at any of the cells, making it able to detect several signals at once—the same way a huge satellite dish can detect more signals than a small dish.
Electrical synapses likely evolved because of evolutionary pressures that selected for speed. These synapses can pass signals as fast as electrical charges can move through an electrolyte-rich fluid like cytoplasm; almost instantaneous.
Another advantage of electrical synapses is that they can form a large network of interconnected neurons with synchronized activity. For example, neuroendocrine cells in the hypothalamus are connected by electrical synapses. When the “go” signal arrives, all the cells depolarize at once, which can result in the massive release of hormones into the bloodstream. A network can also cause sudden, powerful inhibition. Like an angry mob of people chanting, a network of electrical synapses connecting inhibitory interneurons allows the network to send an immediate “shutdown” signal under specific circumstances.
Animation 11.2. Since an electrical synapse is a direct, physical connection between two neurons, ions are able to flow either direction across the gap junction. ‘Bidirectional Electrical Synapse’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC BY-NC-SA) 4.0 International License. View static image of animation.
Electrical synapses share the cytoplasm between the two connected cells, so ions, ATP, and larger signaling molecules and proteins are able to move between the two cells. These signaling molecules play an important role in cellular mechanisms, which we will see in a later chapter.
Animation 11.3. Gap junctions are large enough to allow the flow of small cellular molecules like ATP or second messengers. ‘Electrical Synapse – Small Molecules’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC BY-NC-SA) 4.0 International License. View static image of animation.
Chemical
At a chemical synapse, a signaling molecule is released by the presynaptic cell to influence the postsynaptic cell by binding to postsynaptic receptors. Since chemical synapses do not rely on a direct physical protein “tunnel” to connect the two neurons, the distance between the two cells can be much larger.
On average, a chemical synapse is a distance of about 20-40 nanometers, roughly a thousand times smaller than the diameter of a human hair. A chemical synapse can pass a variety of signals, depending on the neurotransmitter and the receptor. For example, some signals are directly excitatory and allow positively charged cations to enter the neuron causing depolarization. Other signals are hyperpolarizing, and therefore inhibitory. And yet other signals are much more complex, inducing changes in protein expression that can modify cellular excitability over the course of minutes or hours.
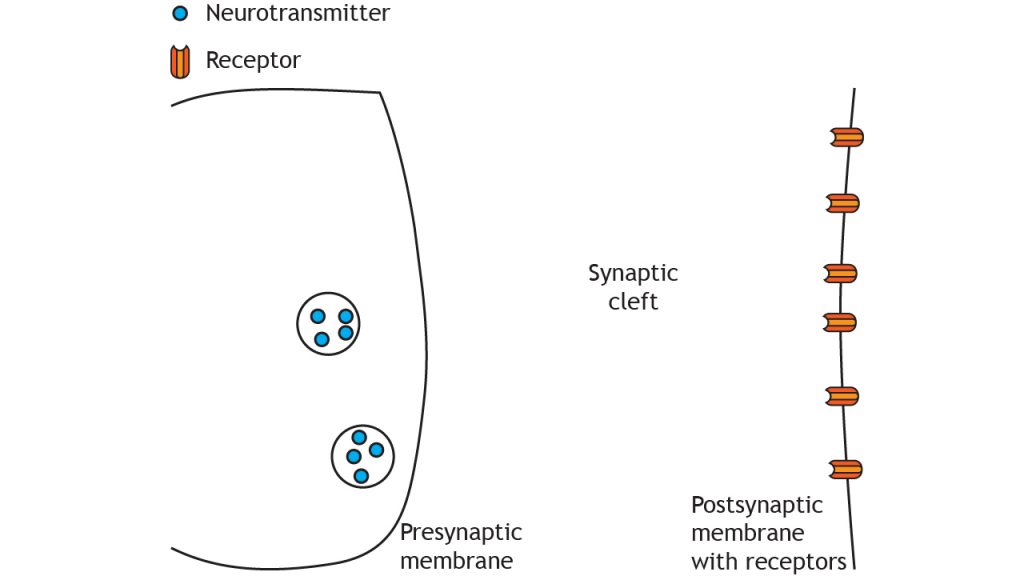
At a chemical synapse, the depolarization of an action potential reaching the presynaptic terminal causes release of neurotransmitters. These neurotransmitters are synthesized and stored in neurons. After being released, these neurotransmitters diffuse randomly across the synapse, where they are able to affect nearby neurons once the chemical binds to its corresponding receptor located in the cell membrane of the postsynaptic neuron. The structure and function of chemical synapses make them slower than electrical synapses and permit signaling in only one direction.
Because of the complexity of the signals that chemical synapses can convey, evolutionary development through time has allowed for a tremendous variety of responses. Chemical synapses allow for fine-tuning of neural networks, giving these nervous systems a larger range of possibilities. The nervous systems of “higher” organisms like humans tend to have several chemical synapses since these signals are likely necessary for complex behaviors and cognition.
Animation 11.4. An action potential causes release of neurotransmitters from the presynaptic terminal into the synaptic cleft. The transmitters then act on neurotransmitter receptors in the postsynaptic membrane. ‘Chemical Synapse – Neurotransmitter Release’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC BY-NC-SA) 4.0 International License. View static image of animation.
Synapse Location
As we discuss synaptic transmission, we will focus mainly on axodendritic synapses, in which the presynaptic terminal (axon) synapses on the dendrites of the postsynaptic cell. But synapses can also be located between the terminal and the cell body of the postsynaptic cell, called axosomatic, or even between the terminal and the axon of the postsynaptic cell, called axoaxonic.

Key Takeaways
- Electrical synapses make direct contact between neurons, are faster than chemical synapses, and can be bidirectional
- Chemical synapses form a synaptic cleft between the neurons and are unidirectional
- Synapses can occur between the presynaptic terminal and the postsynaptic dendrites (axodendritic), cell body (axosomatic), or axon (axoaxonic)
Test Yourself!
An interactive H5P element has been excluded from this version of the text. You can view it online here:
https://openbooks.lib.msu.edu/introneuroscience1/?p=73#h5p-9
Attributions
Portions of this chapter were remixed and revised from the following sources:
- Foundations of Neuroscience by Casey Henley. The original work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License
- Open Neuroscience Initiative by Austin Lim. The original work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.