
3.2: Steps in Synaptic Signaling
- Page ID
- 69848
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Glossary Terms
- Key Takeaways
- Test Yourself
Steps in Chemical Signaling
As we have covered, when an action potential propagates down the axon to the presynaptic terminal, the electrical signal will result in a release of chemical neurotransmitters that will communicate with the postsynaptic cell.
At a chemical synapse, the process of neurotransmitter release is very tightly regulated. If there were no mechanisms to control the release of chemicals at the synapse, nerve cells would deplete their entire stock of neurotransmitter. Regulation of release depends on several proteins that are important parts of the process. These proteins are often embedded within cell membranes of the vesicles or the neuronal membrane.
Step 1: An action potential arrives at the axon terminal
There are a series of steps that take place during chemical synaptic transmission.
First, an action potential propagates down the axon until it arrives at the axon terminal, depolarizing the membrane of the presynaptic terminal.
Animation 12.1. The action potential is a brief, but significant, change in electrical potential across the membrane. The membrane potential will move from a negative, resting membrane potential (shown here as -65 mV) and will rapidly become positive and then rapidly return to rest during an action potential. The action potential moves down the axon beginning at the axon hillock. When it reaches the synaptic terminal, it causes the release of chemical neurotransmitter. ‘Action Potential Propagation’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation
Step 2: Membrane depolarization from action potential causes influx of calcium ions
When the action potential reaches the terminal, there is an influx of sodium ions. This inward positive current causes a depolarization of the terminal, activating voltage-gated calcium channels that are embedded in the cell membrane of the axon terminals. Due to the electrochemical gradient of calcium, when the voltage-gated calcium channels are opened, calcium will rush into the cell. The concentration of intracellular calcium, generally in the range of 100 nM, is much lower than the concentration outside the cell, therefore there is a strong electrochemical gradient that moves calcium into the terminal. As it turns out, an elevation of Ca2+ in the intracellular space is the “go ahead” signal that causes neurotransmitter release.
Animation 12.2. An action potential causes an influx of sodium in the terminal. The depolarization opens voltage-gated calcium channels, and calcium ions flow into the terminal down their electrochemical gradient. The blue, dotted channels represent voltage-gated sodium channels, and the purple, striped channels represent voltage-gated calcium channels. ‘Terminal Calcium Influx’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
Active Zones
The voltage-gated calcium channels are concentrated in the presynaptic terminal at active zones, the regions of the membrane where small molecule neurotransmitters are released. At active zones, some synaptic vesicles are docked and are ready for immediate release upon arrival of the action potential. Other neurotransmitter-filled vesicles remain in a reserve pool outside of the active zone.
Vesicles filled with neuropeptides do not dock at active zones. They are located outside of the active zone, further away from the membrane and the high density of voltage-gated calcium channels. They are, therefore, slower to release than the small molecule transmitters.

Location of vesicles
Synaptic vesicles can be found in one of three places at the axon terminal.
- Readily releasable pool. These vesicles are located close to the cell membrane at the axon terminal. In fact, many of them are already “docked”, meaning that their coat proteins are already interacting closely with the proteins on the inside of the cell membrane. When the depolarizing charge of an action potential reaches the terminal, these vesicles at the readily releasable pool are the first ones that fuse with the cell membrane and release their contents.
- Recycling pool. These vesicles are the ones that have been depleted due to release. They are currently in the process of being refilled or reloaded with neurotransmitter. They are farther from the cell membrane, and the protein machinery is not primed for release, so it requires a more intense stimulation to release the contents of these vesicles.
- Reserve pool. These vesicles are the farthest from the surface of the cell membrane, and most vesicles are held in this reserve pool. For these neurotransmitters to be released, very intense stimulation is required.
Step 3: Docking of synaptic vesicles at the membrane
Docking of synaptic vesicles packaged with small molecule neurotransmitters occurs through the interaction of three membrane-bound proteins called SNARE proteins. Synaptobrevin is called a v-SNARE because it is located on the Vesicular membrane. Syntaxin and SNAP-25 are called t-SNARES because they are located on the terminal membrane, which is the Target membrane. The interaction of these three proteins leads to vesicle docking at the active zone.

The influx of calcium through the voltage-gated calcium channels initiates the exocytosis process that leads to neurotransmitter release. Calcium enters the cell and interacts with a vesicle-bound protein called synaptotagmin. Synaptotagmin is a calcium sensor that detects elevated levels of calcium in the axon terminal.
In the presences of calcium, the v-SNAREs and the t-SNAREs interact with one another, forming a molecular structure called a SNARE complex. The SNARE complex looks a lot like two twist ties that are wound tightly together. As they twist tighter together, it causes the vesicle membrane to approach the inside of the cell membrane, which results in vesicular fusion.
Animation 12.3. Calcium enters the cell when the voltage-gated channels open. In the presence of calcium, synaptotagmin, a protein bound to the vesicular membrane interacts with the SNARE proteins. The purple, striped channels represent voltage-gated calcium channels. ‘Synaptotagmin’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation
Step 4: Release of neurotransmitters into the synapse
The last step of neurotransmitter release is the fusing of the cell membrane. In order to release their chemical contents into the synapse, vesicles need to fuse with the cell membrane. As the vesicular membrane merges with the interior of the neuronal membrane, the membranes fuse and the contents of the vesicle become exposed to the extracellular space. The neurotransmitters then float across the aqueous synapse, giving them the opportunity to interact with postsynaptic receptors.
Animation 12.4. Once the synaptotagmin-SNARE protein complex forms, the synaptic vesicle membrane fuses with the terminal membrane, and the neurotransmitters are released into the synaptic cleft through exocytosis. The purple, striped channels represent voltage-gated calcium channels. ‘Transmitter Exocytosis’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
Step 5: Receptor Activation
After exocytosis of the transmitter molecules, neurotransmitters traverse across the synapse and bind to receptors on the postsynaptic membrane. Receptors fall into two main categories: ionotropic receptors (also called ligand-gated channels) and metabotropic receptors (also called G-protein coupled receptors). These two types of receptors will be covered in upcoming chapters.
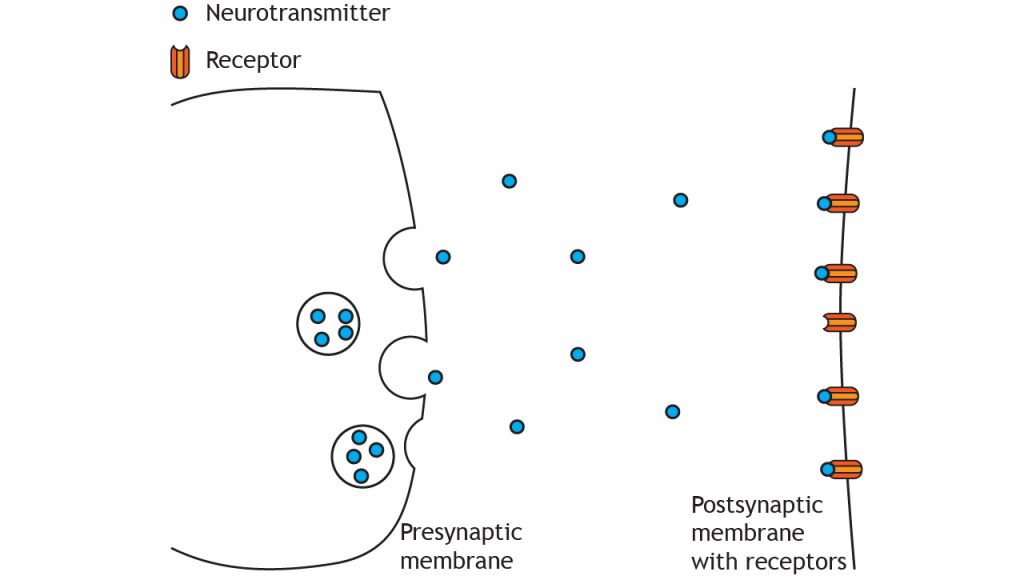
Step 6: Postsynaptic potentials generated in postsynaptic cell summate
Recall the process of summation from Chapter 8. The postsynaptic potentials generated from neurotransmitter binding summate at the axon hillock. If the membrane potential is over threshold potential for the cell, then a new action potential will be generated in the postsynaptic cell.
Postsynaptic potentials (graded potentials) can occur following different types of stimulation.
When neurotransmitters bind to receptors on postsynaptic cells, they cause the receptor proteins to undergo a conformational change, and open ligand-gated ion channels that produce depolarization or hyperpolarization.

Step 7: Neurotransmitter Inactivation
It is important that neurotransmitter signaling be tightly regulated. This means that there needs to be ways to terminate chemical signaling. Inactivation can be accomplished in three different ways.
- After being released, neurotransmitters can be altered into inactive substances.
- Neurotransmitters can go through the reuptake process, where they are recycled by being transported back into the presynaptic cell and repackaged into synaptic vesicles.
- Some neurotransmitters simply float away from the synapse due to the aqueous environment surrounding neurons.
Key Takeaways
- In order for neurotransmitter release to be initiated, an action potential (depolarizing stimulus) needs to arrive at the axon terminal to open voltage-gated calcium channels
- Neurotransmitter release is dependent on the influx of calcium into the terminal through voltage-gated calcium channels
- SNARE proteins are important for vesicle docking at active zones and exocytosis of neurotransmitters into the synapse
- Synaptotagmin is a calcium sensor
- Neurotransmitters bind to postsynaptic receptors and cause postsynaptic potentials that summate within the postsynaptic cell
- Neurotransmitter signaling is terminated through chemical inactivation, reuptake, or through diffusion away from the synapse.
Test Yourself!
An interactive H5P element has been excluded from this version of the text. You can view it online here:
https://openbooks.lib.msu.edu/introneuroscience1/?p=92#h5p-26
Attributions
Portions of this chapter were remixed and revised from the following sources:
- Foundations of Neuroscience by Casey Henley. The original work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License
- Open Neuroscience Initiative by Austin Lim. The original work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.