
5.6: Vestibular System
- Page ID
- 69871
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Glossary Terms
- Key Takeaways
- Test Yourself
When we tilt our head to the side, or look up and down, that movement information is conveyed to our brain using the vestibular system. The vestibular system is a sort of three-dimensional compass that can detect head movement, and that information helps us figure out how our head is oriented and how to balance ourselves in changing conditions. The vestibular system is made up of two structures that are intimately tied in with the anatomical features of the inner ear.
Adjacent to the cochlea within the inner ear is a structure called the vestibular labyrinth made up of the otolith organs (saccule and utricle) and the semicircular canals.
Otolith Organs
Next to the cochlea and within the vestibular labyrinth are two membranous sacs, the saccule and the utricle. Collectively, these structures are called the otolith organs and are responsible for determining gravity through the tilt of the head and linear acceleration. These structures are centrally located within the vestibular labyrinth.
Due to their differences in structure, the two otolith organs have slightly different functions. The saccule is more sensitive to vertical movements, like when you are standing in a moving elevator. The utricle is more responsive to horizontal movements, such as when driving.

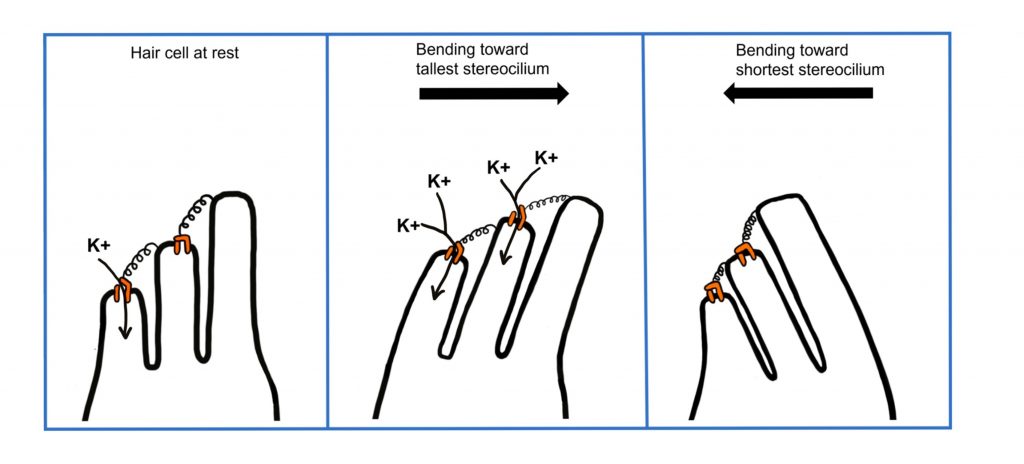
Similar to the other inner ear structure, the cochlea, the structures of the vestibular labyrinth also contain hair cells. The hair cells located within the otolith organs are biologically similar to the hair cells in the cochlea. These hair cells have stereocilia that extend into a gelatinous cap that contains otoconia, which are small crystals of calcium carbonate. The otolith organs are important in sensing gravity. Tilting the head causes the otoconia to move (due to gravity), which will push the hair cell stereocilia and cause them to bend.

The tallest stereocilium (singular of stereocilia) is called the kinocilium. Similar to the function of hair cells within the cochlea, movement that bends the stereocilia toward the kinocilium results in depolarization due to potassium influx through open mechanically-gated ion channels within the tips of the stereocilia. Movement that bends the stereocilia away from the kinocilium causes the mechanically-gated ion channels to close, decreasing potassium influx and causing hyperpolarization.
Semicircular Canals
The other major structure of the vestibular labyrinth are the semicircular canals. The semicircular canals are the structures that are responsible for detecting head rotation and angular acceleration.
Anatomically, the semicircular canals are a series of three arch-shaped membranous tubes within the vestibular labyrinth, each one oriented at a right angle to each other. Because of this shape, the semicircular canals sense and convey information about any direction of head movement.
These semicircular canals are filled with endolymph, the same potassium-rich solution that is in the cochlea that is important for auditory sensation. At the end of each of the three canals is a small swelling called the ampulla. Contained in the ampulla is a gelatinous membrane called the cupula. Here, hair cells extend stereocilia into the cupula. The tallest stereocilium is again referred to as the kinocilium.
When we rotate our head, the endolymph within the semicircular canals has a delay in movement due to inertia, and as a result the endolymph will move the cupula in the opposite direction of head movement. The physical movement of the cupula also pushes the cilia of the hair cells within the cupula. As previously discussed in the cochlea and the otolith organs, these hair cells also have mechanically-gated ion channels located in the tips of the stereocilia and function in the same way, with deflection towards the kinocilium resulting in depolarization and deflection away from the kinocilium resulting in hyperpolarization.
When we stop rotating, the endolymph within the semicircular canals continues to move for a brief period of time due to inertia (which contributes to our feeling of dizziness when we stop rotating).

Central Vestibular Pathways
The vestibular system is responsible for coordinating our balance through our head and body movements. As such, the neurons of the vestibular system signal to motor neurons that adjust head, eye, and body position.
The vestibular hair cell information is passed to the brain via excitatory synapses onto a branch of the vestibulocochlear nerve (CN VIII). The cell bodies of these neurons are in Scarpa’s ganglion. These axons send projections to several brain areas, notably the cerebellum, which is a structure critically important for balance. CN VIII also projects into the vestibular nuclei of the medulla. From there, neurons signal to areas of the thalamus that ultimately control motor areas of the face, extraocular motor neurons that ultimately control eye movement, and the spinal cord that is responsible for limb and neck motor movements to help maintain body and head position.

Key Takeaways
- The vestibular labyrinth is located within the inner ear and functions in our sense of balance.
- The structures of the vestibular labyrinth are the otolith organs (the saccule and utricle) and the three semicircular canals.
- The vestibular labyrinth structures also contain hair cells similar to the hair cells within the auditory system. Bending the stereocilia in the direction of the tallest stereocilium results in depolarization of the hair cell and bending in the direction of the shortest stereocilium results in hyperpolarization.
- The vestibular labyrinth structures signal to multiple brain areas to control head movement, eye movements, and balance.
Test Yourself
An interactive H5P element has been excluded from this version of the text. You can view it online here:
https://openbooks.lib.msu.edu/introneuroscience1/?p=647#h5p-40
Attributions
Portions of this chapter were remixed and revised from the following sources:
- Open Neuroscience Initiative by Austin Lim. The original work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
Media Attributions
- Vestibular organs © NASA adapted by Valerie Hedges is licensed under a Public Domain license
- Otolith organ hair cells © Valerie Hedges is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Hair cell depolarization and hyperpolarization © Valerie Hedges is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Semicircular canal cupula © NASA adapted by Valerie Hedges is licensed under a Public Domain license
- Central Vestibular Pathway © Valerie Hedges is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license