
5.10: Touch- The Skin
- Page ID
- 69994
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Glossary terms
- Key Takeaways
- Test Yourself
- Additional Review
Touch can come in many forms: pressure, vibration, stretch, motion, edges, points, etc. Receptors in the skin allow for perception of these different characteristics, and when this information is combined in the central nervous system, we are able to determine the location, strength, duration, movement, shape, and texture of the object interacting with the skin.
Cutaneous (meaning found in the skin) mechanoreceptors are responsible for sensing mechanical changes to the skin.
Receptors
We can feel different modalities of touch because of the presence of specialized sensory receptors, called mechanoreceptors, located in the skin.
There are four different classes of cutaneous mechanoreceptors. These different mechanoreceptors have slightly different shapes and are therefore capable of detecting different types of stimuli.
- Merkel’s discs are located at the border between the dermis and epidermis and are specialized to detect edges and points. They are very densely populated underneath our fingertips. These are the most precise mechanoreceptors with resolution at the level of 0.5 mm, which allow us to detect different distances in bumps while running your fingers across Braille.
- Pacinian corpuscles are mechanoreceptors wrapped in several layers of connective tissue. The Pacinian corpuscles are located deep in the dermis of the skin and are responsible for perception of vibrations.
- Meissner’s corpuscles are located in the epidermis layer and are highly concentrated at the fingertips. They are stimulated by skin motion/movement.
- Ruffini endings are located in the dermis layer of the skin and respond to stretching of the skin.
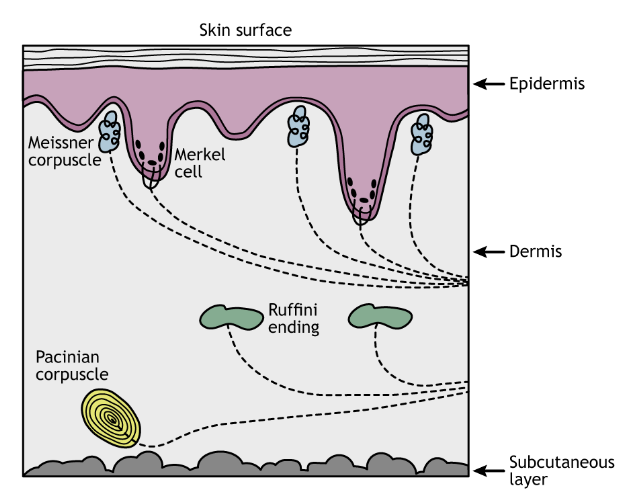
Structure of a Cutaneous Mechanoreceptor
The somatosensory afferents from the cutaneous mechanoreceptor have their cell bodies located in the dorsal root ganglion (DRG), a clump of somata that are close to the dorsal side of the spinal cord. These touch receptors are the primary neurons and are classified as pseudounipolar neurons based on their morphology. Pseudounipolar neurons don’t have true dendrites; instead each soma sends a single protrusion which branches into two directions. One branch goes towards the skin surface where it terminates as one of the mechanoreceptors. Typically, the terminal of this branch is surrounded by a connective tissue covering. The other branch off the soma travels towards the spinal cord.
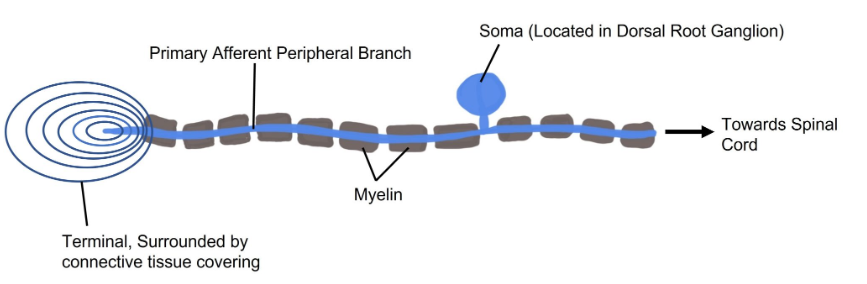
Sensory Transduction
On a molecular scale, these mechanoreceptors detect changes at the skin using mechanically-gated ion channels. In previous chapters we discussed ion channels that are gated by voltage changes and channels that are gated by neurotransmitters. In the somatosensory system, we find ion channels that are gated by physical distortion or stretch of the membrane. These transmembrane proteins are specialized for detecting the physical distortion of the channel and can open by stretch of the membrane itself or indirectly through movement of intra- or extracellular proteins that are linked to the channels. They are specialized for detecting the physical distortion of the channel, similar to the channels found in the stereocilia of the hair cells or vestibular cells.
When pressure is applied to these proteins, the cation channel opens and sodium and calcium flow down their electrochemical gradient into the neuron, causing both a depolarization and the initiation of second messenger cascades. This depolarization is referred to as a ‘receptor potential’. If enough stimulus is applied, the depolarization reaches threshold potential for the cell and an action potential is sent toward the spinal cord.
Animation 37.1 Mechanoreceptors respond to touch stimuli via stretch-gated non-selective cation channels. The channels can either open due to stretch of the membrane itself which stretches open the channel or due to proteins associated with the channels that pull the channel open. ‘Stretch-Gated Ion Channels’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC BY-NC-SA) 4.0 International License. View static image of animation.
Receptive Fields
Each cutaneous mechanoreceptor is able to detect somatosensory information in an area of skin called the receptive field. When the receptive field is touched, the mechanoreceptor will be activated. The size of a receptive field can be determined by measuring the rate of action potentials from the nerve that collects somatosensory information from that patch of skin. The smaller the receptive field, the better the brain can distinguish between two different adjacent tactile stimuli.
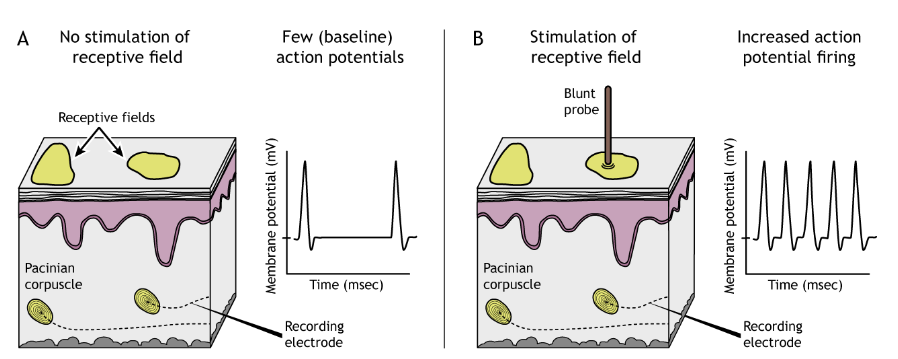
Receptive Field Size
Additionally, different classes of mechanoreceptors have different sizes of receptive fields: deeper structures, like Pacinian corpuscles, have larger receptive fields compared to receptors found closer to the skin surface that have smaller receptive fields. Merkel cells and Meissner corpuscles, both of which are located near the skin surface, have small receptive fields and thus collect information from a restricted area of sensory space. Ruffini endings and Pacinian corpuscles, located deeper in the skin layers, have larger receptive fields than the Merkel cells and Meissner corpuscles and thus collect information over a large are of sensory space.
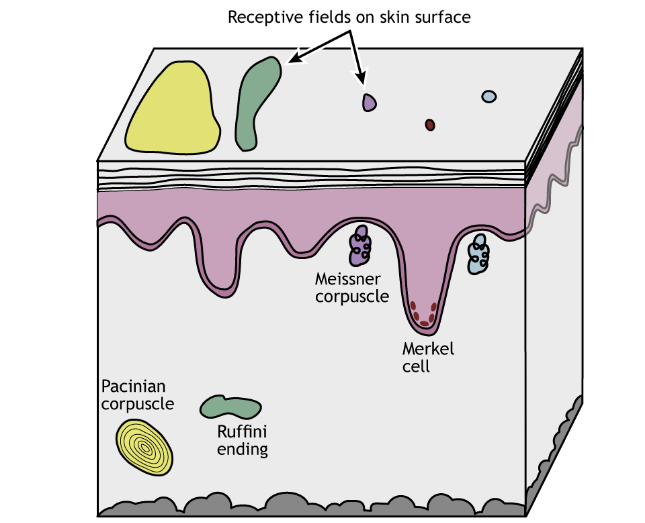
Receptive field sizes are different among the different mechanoreceptors, but they also vary among different body regions. Even within one receptor type (e.g. Meissner corpuscles), receptive fields in regions like the fingers or lips are smaller than in regions like the back or leg. This allows us to have finer spatial resolution with locating and identifying objects using our fingers. The smaller receptive fields in these regions are a result of a higher density of receptors in the skin.
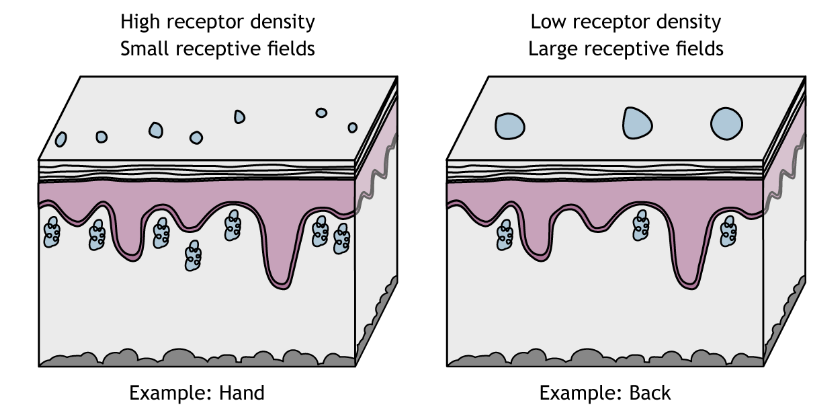
Two-Point Discrimination
Receptive field sizes are important because they allow us to locate a stimulus on our bodies. Larger receptive fields are not as precise as smaller receptive fields. One measure of receptive field size is two-point discrimination (try it at home!), which determines the minimum distance needed between two stimuli to perceive two separate points on the skin and not one. The hand has a smaller threshold for discerning between two points than does the back, a result of the different sized in receptive fields. In skin areas with a large receptive field, two adjacent stimuli may feel like one.
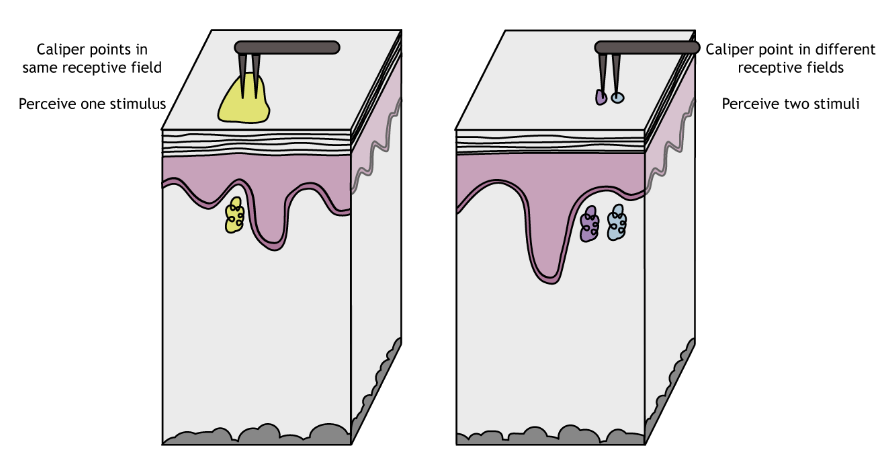
Skin areas with small receptive fields are better able to distinguish two similarly-spaced points. The hand has a smaller threshold for discerning between two points than does the back, a result of the different sized in receptive fields.
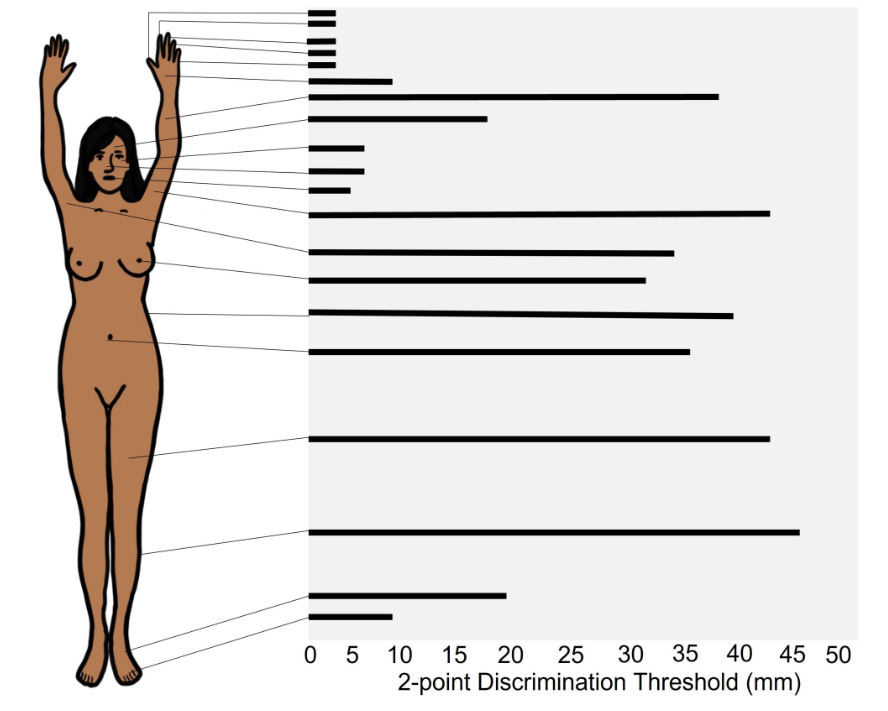
The two-point discrimination threshold for the fingers is very small, which indicates that they have high sensitivity. Body areas with high sensitivity typically have higher density of mechanoreceptors, have receptors that have smaller receptive fields, and have more brain tissue devoted to processing the sensory information that comes from those structures.
The two-point discrimination threshold for areas such as the leg or torso is very large, which indicates that these areas have low sensitivity. Body areas with low sensitive have a lower density of mechanoreceptors, have receptors that have larger receptive fields, and have less brain tissue devoted to sensory information that comes from those structures.
Adaptation Rate
Another important characteristic of the somatic sensory receptors is that of adaptation rate. The class of mechanoreceptors can be divided into two categories based on their electrical properties in response to stimuli: slowly adapting and rapidly-adapting.
Slowly-adapting mechanoreceptors change their action potential firing rate as long as the stimulus is present, while rapidly-adapting mechanoreceptors only change activity at the moment there is a change in stimulus. For example, imagine the stimulus of a coin that is sitting in the palm of your hand. The slowly-adapting sensory neurons may increase their firing rate as long as the coin is in your palm. When the quarter is removed, they return to their baseline firing rate. Slowly-adapting mechanoreceptors change their action potential firing rate as long as the stimulus is present and are most useful for determining the pressure and shape of a stimulus. Merkel cells and Ruffini endings are both slowly adapting fibers.
Animation 37.2. Slowly adapting mechanoreceptors continuing firing action potentials throughout the duration of a stimulus. As the stimulus moves from not present, to weak, to strong, the action potential firing of the Ruffini ending fires throughout the entire stimulus. ‘Slowly Adapting Receptor’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC BY-NC-SA) 4.0 International License. View static image of animation.
Rapidly-adapting mechanoreceptors only change activity at the moment there is a change in stimulus, but not when a stimulus is constant. This firing makes rapidly adapting fibers specialized for detecting movement and vibration. Considering the coin example from above, a rapidly adapting sensory neuron will change its action potential firing properties only at two timepoints: the moment the coin lands in your hand, and the moment that coin is removed. These neurons signal a change in status.
Animation 37.3 Rapidly adapting mechanoreceptors firing action potentials when the strength of the stimulus changes. As the stimulus moves from not present, to weak, to strong, the action potential firing of the Pacinian corpuscle only fires when the stimulus changes strength. ‘Rapidly Adapting Receptor’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC BY-NC-SA) 4.0 International License. View static image of animation.
Key Takeaways
- There are multiple types of mechanoreceptors in the skin that are activated by different types of touch stimuli
- The receptive field size differs among the types of mechanoreceptors
- The adaptation rate differs among the types of mechanoreceptors
- Receptive field is a region of skin that activate a given mechanoreceptor
- Receptive field size for a specific type of mechanoreceptor can vary in size across the body
- Mechanoreceptors express stretch-gated non-selective ion channels that depolarize the cell during sensory transduction
Test Yourself!
Additional Review
Describe the relationship between density of receptors, receptive fields, and two-point discrimination.
Attributions
- Foundations of Neuroscience by Casey Henley. The original work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License
- Open Neuroscience Initiative by Austin Lim. The original work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
Media Attributions
- Mechanoreceptors © Casey Henley is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- somatosensory afferent © Valerie Hedges is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Receptive Field Activation © Casey Henley is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Mechanoreceptor Receptive Fields © Casey Henley is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Receptive Field Location © Casey Henley is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Two Point Discrimination © Casey Henley is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- 2 point discrimination © Valerie Hedges is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license