
9.3: Skeletal Muscle
- Page ID
- 22317
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Describe the layers of connective tissues that package skeletal muscle
- Explain how muscles work with tendons to move the body
- Identify and describe the microscopic anatomy of a muscle fiber
- Describe excitation-contraction coupling
The best-known feature of skeletal muscle is its ability to contract and cause movement. Skeletal muscles act not only to produce movement but also to stop movement, such as resisting gravity to maintain posture. Small, constant adjustments of skeletal muscles are needed to hold a body upright or balanced in any position. Muscles also prevent excess movement of the bones and joints, maintaining skeletal stability and preventing structural damage or deformation. Joints can become misaligned or dislocated by excessive or inappropriate forces applied to associated bones; muscles work to keep joints stable. Skeletal muscles are located throughout the body at the openings of internal tracts to control the movement of various substances. These muscles allow functions, such as swallowing, urination, and defecation, to be under voluntary control. Skeletal muscles also protect internal organs (particularly abdominal and pelvic organs) by acting as an external barrier or shield to external trauma and by supporting the weight of the organs.
Skeletal muscles contribute to the maintenance of homeostasis in the body by generating heat. Muscle contraction requires energy, and when ATP is broken down, heat is produced. This heat is very noticeable during exercise, when sustained muscle movement causes body temperature to rise, and in cases of extreme cold, when shivering produces random skeletal muscle contractions to generate heat.
Each skeletal muscle is an organ that consists of various integrated tissues. These tissues include the skeletal muscle fibers, blood vessels, nerve fibers, and connective tissue. Each skeletal muscle has three layers of connective tissue (called “mysia”) that enclose it and provide structure to the muscle as a whole, and also compartmentalize the muscle fibers within the muscle (Figure \(\PageIndex{1}\)). Each muscle is wrapped in a sheath of dense, irregular connective tissue called the epimysium, which allows a muscle to contract and move powerfully while maintaining its structural integrity. The epimysium also separates muscle from other tissues and organs in the area, allowing the muscle to move independently.
Inside each skeletal muscle, muscle fibers are organized into bundles, called a fascicle, by a middle layer of dense irregular connective tissue called the perimysium. This fascicular organization is common in muscles of the limbs; it allows the nervous system to trigger a specific movement of a muscle by activating a subset of muscle fibers within a bundle, or fascicle, of the muscle. Inside each fascicle, each muscle fiber is encased in a thin areolar connective tissue layer of collagen and reticular fibers called the endomysium. The endomysium contains the extracellular fluid and nutrients to support the muscle fiber. These nutrients are supplied via blood capillaries located in the endomysium.
Figure \(\PageIndex{1}\): The Three Connective Tissue Layers. Bundles of muscle fibers, called fascicles, are covered by the perimysium. Muscle fibers are covered by the endomysium. (Image credit: "The Three Connective Tissue Layers" by Openstax is licensed under CC BY 4.0)
In skeletal muscles that work with tendons to pull on bones, the collagen in the three tissue layers intertwines and integrates with the collagen of a tendon, straightening to form a cord of dense regular connective tissue. At the other end of the tendon, the collagen incorporates into periosteum becoming dense irregular connective tissue once more. Tension created by contraction of the muscle fibers is transferred though the mysia, to the tendon, and then to the periosteum to pull on the bone to move the skeleton. In other places, the mysia may fuse with a broad, tendon-like sheet called an aponeurosis, or to fascia, the connective tissue between skin and bones. The broad sheet of connective tissue in the lower back that the latissimus dorsi muscles (the “lats”) fuse into is an example of an aponeurosis.
Every skeletal muscle is richly supplied by blood vessels for nourishment, oxygen delivery, and waste removal. In addition, every muscle fiber in a skeletal muscle is supplied by the axon branch of a somatic motor neuron, which signals the fiber to contract. Unlike cardiac and smooth muscle, the only way to functionally contract a skeletal muscle is through signaling from the nervous system.
The Neuromuscular Junction
The specialized site of the skeletal muscle where a motor neuron’s axon terminal meets the muscle fiber is called the neuromuscular junction (NMJ) (Figure \(\PageIndex{2}\)). This is where the muscle fiber first responds to signaling by the motor neuron. The sarcolemma (plasma membrane) in the region of a neuromuscular junction, known as the motor end-plate, contains special transmembrane proteins that receive a signal from the motor neuron and begin the electrical signal in the muscle fiber (Figure \(\PageIndex{5}\)). Every skeletal muscle fiber in every skeletal muscle is innervated by a motor neuron at the NMJ. Excitation signals from the neuron are the only way to functionally activate the fiber to contract.
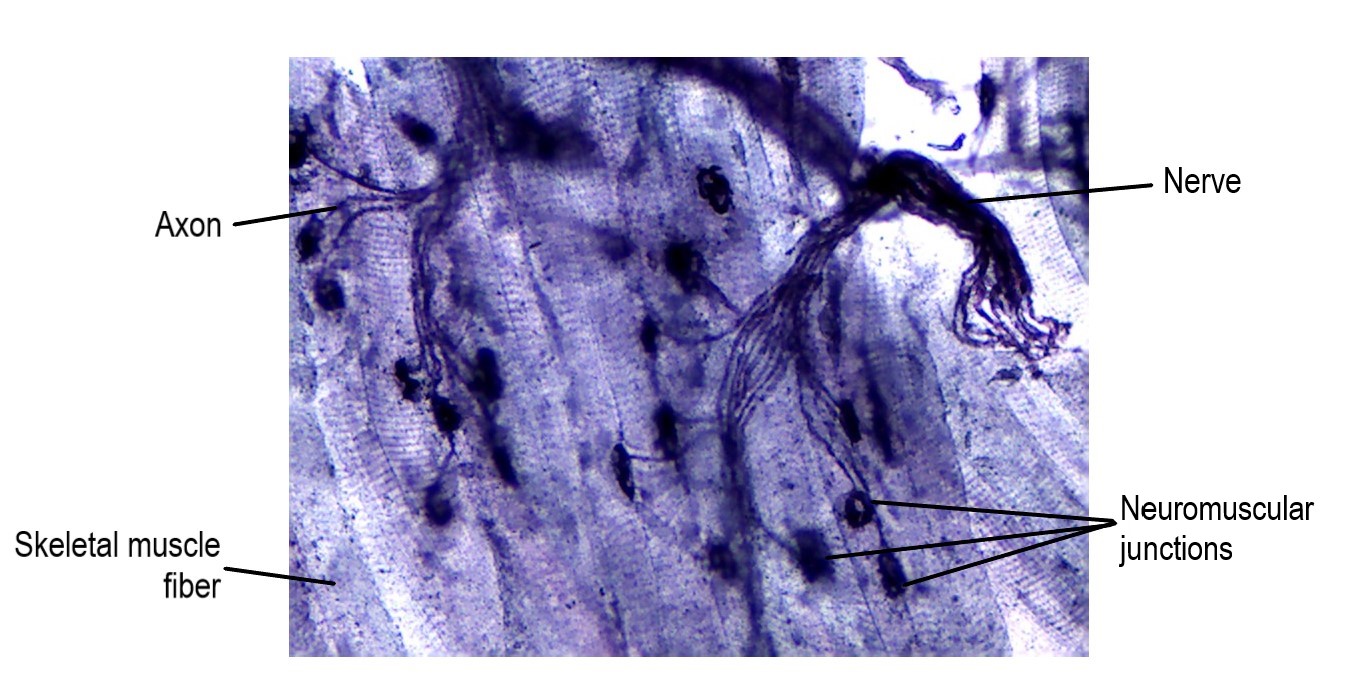
Skeletal Muscle Fibers
Because skeletal muscle cells are long and cylindrical, they are commonly referred to as skeletal muscle fibers. Specific terminology associated with muscle fibers is rooted in the Greek sarco, which means “flesh.” The plasma membrane of muscle fibers is called the sarcolemma and the cytoplasm is referred to as sarcoplasm. Skeletal muscle fibers can be quite large for human cells, with diameters up to 100 μm and lengths up to 30 cm (11.8 in). During early development, embryonic myoblasts, each with its own nucleus, fuse with up to hundreds of other myoblasts to form the multinucleated skeletal muscle fibers. Multiple nuclei mean multiple copies of genes, permitting the production of the large amounts of proteins and enzymes needed for muscle contraction along the length of the entire muscle fiber.
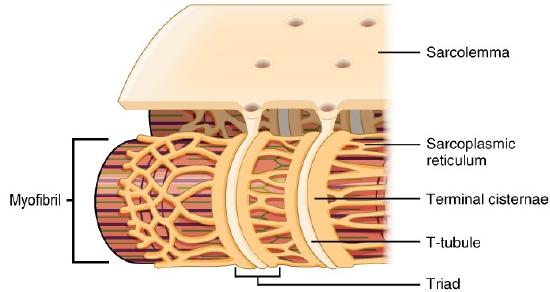
Within the sarcoplasm of each muscle fiber are myofibrils that run the entire length of the muscle fiber and attach to the sarcolemma at its end (Figure \(\PageIndex{1}\)). Each myofibril is composed of a bundle of contractile myofilaments (known as actin and myosin), along with other support proteins. As myofibrils contract, the entire muscle cell contracts. Because myofibrils are only approximately 1.2 μm in diameter, hundreds to thousands can be found inside one muscle fiber. Surrounding each myofibril is the sarcoplasmic reticulum (SR), the specialized smooth endoplasmic reticulum of muscle fibers, which stores, releases, and retrieves calcium ions (Ca++) (Figure \(\PageIndex{3}\)).
As will be discussed, the signal from a motor neuron actually stimulates the release of calcium ions (Ca++) from its storage in the cell’s SR. For the action potential to reach the membrane of the SR from the motor end plate, there are periodic invaginations in the sarcolemma, called T-tubules (“T” stands for “transverse”). You will recall that the diameter of a muscle fiber can be up to 100 μm, so these T-tubules ensure that the membrane can get close to the SR in the sarcoplasm. On each side of the T-tubule is a terminal cisterna (plural - cisternae), an enlarged region of the SR that contains high concentrations of calcium ions. The arrangement of a T-tubule with the terminal cisternae of the SR on either side is called a triad (Figure \(\PageIndex{3}\)). The triad surrounds the cylindrical structure of the myofibril.
The Sarcomere
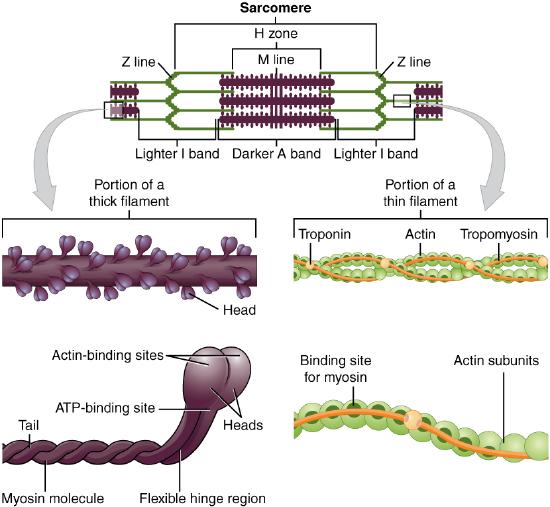
The striated appearance of skeletal muscle fibers is due to the arrangement of the myofilaments actin and myosin in sequential order from one end of the muscle fiber to the other. Each segment of these myofilaments and their regulatory proteins, troponin and tropomyosin (along with other proteins), is called a sarcomere. The sarcomere is the functional unit of the muscle fiber. A chain of thousands of repeating sarcomeres forms the long tube-like myofibrils.
Each sarcomere is approximately 2 μm in length with a three-dimensional cylinder-like arrangement. The repeating pattern of the myofilaments, along with other proteins, within the sarcomere gives rise to very distinctive regions and structures (Figure \(\PageIndex{4}\)). Each sarcomere is bordered by structures called Z-discs (also called Z-lines, because pictures are two-dimensional), to which the actin myofilaments are anchored. Because the actin forms strands that are thinner than the myosin, it is called the thin filament of the sarcomere. Likewise, because the myosin strands have more mass and are thicker, they are called the thick filament of the sarcomere. This difference in thickness, is what causes the striated appearance of skeletal muscle fibers. The area immediately surrounding the Z-discs, is known as the I band, or light band, because only actin is present. The thin composition of actin makes it difficult to see, hence the name the light band. The myosin is anchored in the center of the sarcomere, to a structure known as the M line, where is projects toward but not all to way to, the Z-discs. The area of the sarcomere where myosin is present is known as the A band, or dark band. Opposite to thin composition of actin, myosin is thick, which makes this area dark in appearance. There are some areas, where actin and myosin myofilaments are both present, known as the zone of overlap. Immediately surrounding the M line is the H zone, where there is no overlap and only myosin is present.
Figure \(\PageIndex{4}\): The Sarcomere. Myofibrils within the sarcoplasm are composed of chains of thousands of repeating sarcomeres. (Image credit: "Muscle Fibers" by Openstax is licensed under CC BY 4.0)
Myofilaments
As mentioned above, actin is known as the thin filament and myosin as the thick filament. The structural differences between these two myofilaments not only leads to the striated appearance of skeletal muscle tissue, but also to the ability of the two to work together to produce the overall function of muscle contraction.
Actin myofilaments are formed by multiple actin subunits, each of which contain a myosin binding site (Figure \(\PageIndex{5}\)). In a relaxed muscle, the binding sites remain shielded by the troponin-tropomyosin regulatory complex. The tropomyosin filaments run the length of the actin, covering all of the binding sites and are held in place by troponin.
Myosin myofilaments have two specific regions, the tail and the heads (Figure \(\PageIndex{5}\)). Connecting these two regions is a flexible hinge region. The tails function to cluster many myosin myofilaments together, while the heads take an active role in the muscle contraction process. The heads contain an actin-binding site, as well as an ATP binding site. The importance of these binding sites will be discussed in the next section of this book.
Excitation-Contraction Coupling
All living cells have membrane potentials, or electrical gradients across their membranes. The inside of the membrane is usually around -60 to -90 mV, relative to the outside. This is referred to as a cell’s membrane potential. Neurons and muscle cells can use their membrane potentials to generate electrical signals. They do this by controlling the movement of charged particles, called ions, across their membranes to create electrical currents. This is achieved by opening and closing specialized proteins in the membrane called ion channels. Although the currents generated by ions moving through these channel proteins are very small, they form the basis of both neural signaling and muscle contraction.
Both neurons and skeletal muscle cells are electrically excitable, meaning that they are able to generate action potentials. An action potential is a special type of electrical signal that can travel along a cell membrane as a wave. This allows a signal to be transmitted quickly and faithfully over long distances.
Although the term excitation-contraction coupling confuses or scares some students, it comes down to this: for a skeletal muscle fiber to contract, its membrane must first be “excited”—in other words, it must be stimulated to fire an action potential. The muscle fiber action potential, which sweeps along the sarcolemma as a wave, is “coupled” to the actual contraction through the release of calcium ions (Ca++) from the SR. Once released, the Ca++ interacts with the shielding proteins, forcing them to move aside so that the actin-binding sites are available for attachment by myosin heads. The myosin then pulls the actin filaments toward the center, shortening the muscle fiber.
In skeletal muscle, this sequence begins with signals from the somatic motor division of the nervous system. In other words, the “excitation” step in skeletal muscles is always triggered by signaling from the nervous system (Figure \(\PageIndex{6}\)).
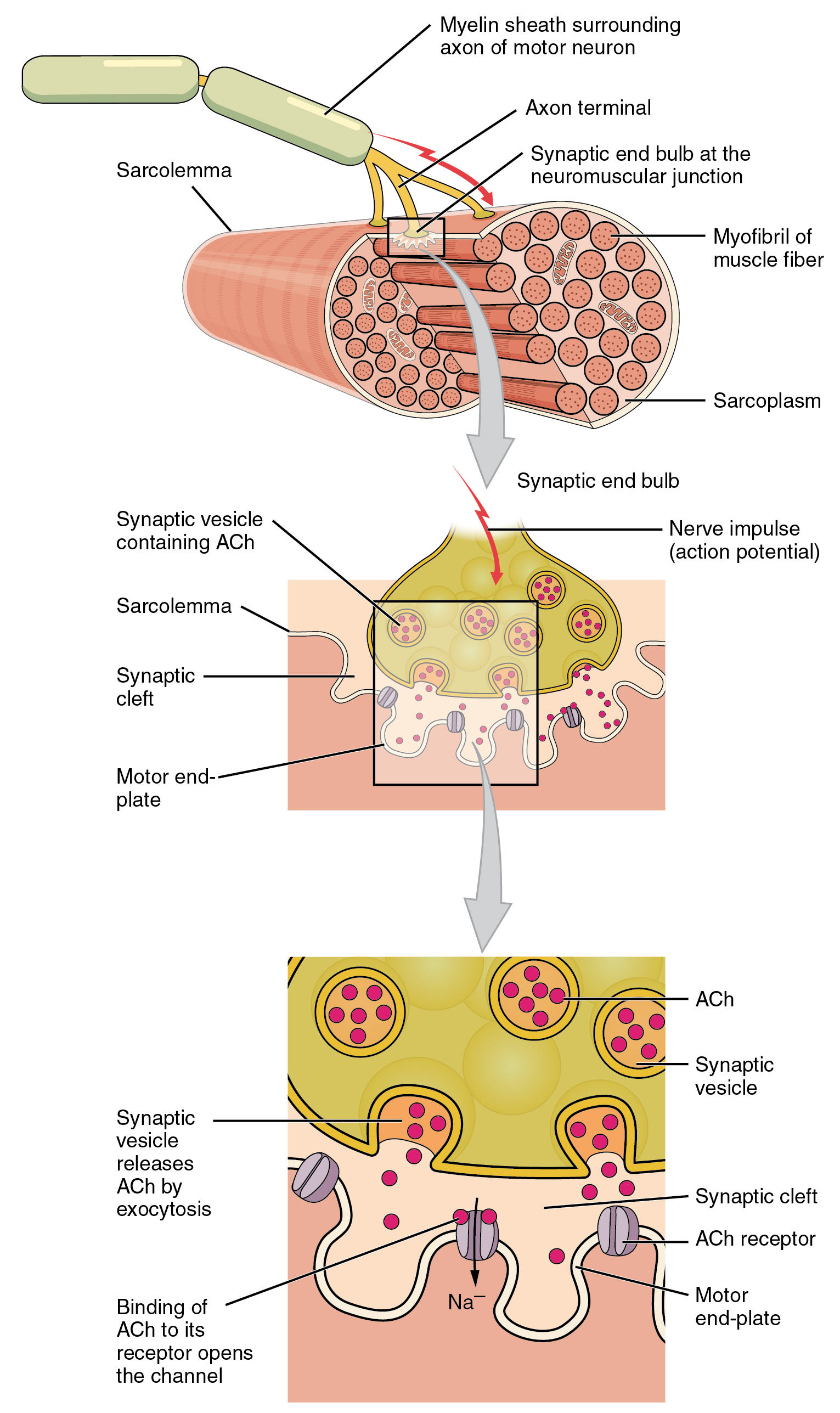
The motor neurons that tell the skeletal muscle fibers to contract originate in the spinal cord, with a smaller number located in the brainstem for activation of skeletal muscles of the face, head, and neck. These neurons have long processes, called axons, which are specialized to transmit action potentials long distances— in this case, all the way from the spinal cord to the muscle itself (which may be up to three feet away). The axons of multiple neurons bundle together to form nerves, like wires bundled together in a cable.
Signaling begins when a neuronal action potential travels along the axon of a motor neuron, and then along the individual branches to terminate at the NMJ. At the NMJ, the axon terminal releases a chemical messenger, or neurotransmitter, called acetylcholine (ACh). The ACh molecules diffuse across a minute space called the synaptic cleft and bind to ACh receptors located within the motor end-plate of the sarcolemma on the other side of the synapse. Once ACh binds, a channel in the ACh receptor opens and positively charged ions can pass through into the muscle fiber, causing it to depolarize, meaning that the membrane potential of the muscle fiber becomes less negative (closer to zero.)
As the membrane depolarizes, another set of ion channels called voltage-gated sodium channels are triggered to open. Sodium ions enter the muscle fiber, and an action potential rapidly spreads (or “fires”) along the entire membrane to initiate excitation-contraction coupling.
Things happen very quickly in the world of excitable membranes (just think about how quickly you can snap your fingers as soon as you decide to do it). Immediately following depolarization of the membrane, it repolarizes, re-establishing the negative membrane potential. Meanwhile, the ACh in the synaptic cleft is degraded by the enzyme acetylcholinesterase (AChE) so that the ACh cannot rebind to a receptor and reopen its channel, which would cause unwanted extended muscle excitation and contraction. Propagation of an action potential along the sarcolemma is the excitation portion of excitation-contraction coupling. The T-tubules carry the action potential into the interior of the cell, which triggers the opening of calcium channels in the membrane of the adjacent SR, causing Ca++ to diffuse out of the SR and into the sarcoplasm. It is the arrival of Ca++ in the sarcoplasm that initiates contraction of the muscle fiber by its contractile units, or sarcomeres.
Concept Review
Skeletal muscles contain connective tissue, blood vessels, and nerves. There are three layers of connective tissue: epimysium, perimysium, and endomysium. Skeletal muscle fibers are organized into groups called fascicles. Blood vessels and nerves enter the connective tissue and branch. Muscles attach to bones directly or through tendons or aponeuroses. Skeletal muscles maintain posture, stabilize joints, support organs, control internal movement, and generate heat.
Skeletal muscle fibers are long, multinucleated cells. The membrane of the cell is the sarcolemma; the cytoplasm of the cell is the sarcoplasm. The sarcoplasmic reticulum (SR) is a form of endoplasmic reticulum that specializes in storing and releasing calcium. Muscle fibers are composed of myofibrils. The banding pattern of myofibrils, called striations, are created by the organization of actin and myosin proteins in functional units called sarcomeres.
Review Questions
Q. The correct order for the smallest to the largest unit of organization in muscle tissue is ________.
A. fascicle, myofilament, muscle fiber, myofibril
B. myofilament, myofibril, muscle fiber, fascicle
C. muscle fiber, fascicle, myofilament, myofibril
D. myofibril, muscle fiber, myofilament, fascicle
- Answer
-
Answer: B
Q. Depolarization of the sarcolemma means ________.
A. the inside of the membrane has become less negative as sodium ions accumulate
B. the outside of the membrane has become less negative as sodium ions accumulate
C. the inside of the membrane has become more negative as sodium ions accumulate
D. the sarcolemma has completely lost any electrical charge
- Answer
-
Answer: A
Critical Thinking Questions
Q. What would happen to skeletal muscle if the epimysium were destroyed?
- Answer
-
A. Muscles would lose their integrity during powerful movements, resulting in muscle damage.
Q. Describe how tendons facilitate body movement.
- Answer
-
A. When a muscle contracts, the force of movement is transmitted through the tendon, which pulls on the bone to produce skeletal movement.
Q. What are the five primary functions of skeletal muscle?
- Answer
-
A. Produce movement of the skeleton, maintain posture and body position, support soft tissues, encircle openings of the digestive, urinary, and other tracts, and maintain body temperature.
Q. What are the opposite roles of voltage-gated sodium channels and voltage-gated potassium channels?
- Answer
-
A. The opening of voltage-gated sodium channels, followed by the influx of Na+, transmits an Action Potential after the membrane has sufficiently depolarized. The delayed opening of potassium channels allows K+ to exit the cell, to repolarize the membrane.
Glossary
- A band
- region of the sarcomere where myosin is present
- acetylcholine (ACh)
- neurotransmitter that binds at a motor end-plate to trigger depolarization
- actin
- protein that makes up most of the thin myofilaments in a sarcomere muscle fiber
- action potential
- change in voltage of a cell membrane in response to a stimulus that results in transmission of an electrical signal; unique to neurons and muscle fibers
- aponeurosis
- broad, tendon-like sheet of connective tissue that attaches a skeletal muscle to another skeletal muscle or to a bone
- depolarize
- to reduce the voltage difference between the inside and outside of a cell’s plasma membrane (the sarcolemma for a muscle fiber), making the inside less negative than at rest
- endomysium
- loose, and well-hydrated connective tissue covering each muscle fiber in a skeletal muscle
- epimysium
- outer layer of connective tissue around a skeletal muscle
- excitation-contraction coupling
- sequence of events from motor neuron signaling to a skeletal muscle fiber to contraction of the fiber’s sarcomeres
- fascicle
- bundle of muscle fibers within a skeletal muscle
- H zone
- region of the sarcomere surrounding the M line where only myosin is present
- I band
- also known as the light band; region of the sarcomere where only actin is present
- M line
- center structure of a sarcomere that serves to anchor myosin filaments
- motor end-plate
- sarcolemma of muscle fiber at the neuromuscular junction, with receptors for the neurotransmitter acetylcholine
- myofibril
- long, cylindrical groups of myofilaments (actin and myosin) that run parallel within the muscle fiber and contains the sarcomeres
- myofilament
- proteins, actin and myosin, that make up myofibrils
- myosin
- protein that makes up most of the thick cylindrical myofilament within a sarcomere muscle fiber
- neuromuscular junction (NMJ)
- synapse between the axon terminal of a motor neuron and the section of the membrane of a muscle fiber with receptors for the acetylcholine released by the terminal
- neurotransmitter
- signaling chemical released by nerve terminals that bind to and activate receptors on target cells
- perimysium
- connective tissue that bundles skeletal muscle fibers into fascicles within a skeletal muscle
- sarcomere
- longitudinally, repeating functional unit of skeletal muscle, with all of the contractile and associated proteins involved in contraction
- sarcolemma
- plasma membrane of a skeletal muscle fiber
- sarcoplasm
- cytoplasm of a muscle cell
- sarcoplasmic reticulum (SR)
- specialized smooth endoplasmic reticulum, which stores, releases, and retrieves Ca++
- skeletal muscle fiber
- multinucleated skeletal muscle cells
- synaptic cleft
- space between a nerve (axon) terminal and a motor end-plate
- T-tubule
- projection of the sarcolemma into the interior of the cell
- terminal cisternae
- enlarged regions of the sarcoplamic reticulum found on either side of the t-tubules
- thick filament
- the thick myosin strands and their multiple heads projecting from the center of the sarcomere toward, but not all to way to, the Z-discs
- thin filament
- thin strands of actin and its troponin-tropomyosin complex projecting from the Z-discs toward the center of the sarcomere
- triad
- the grouping of one T-tubule and two terminal cisternae
- troponin
- regulatory protein that binds to actin, tropomyosin, and calcium
- tropomyosin
- regulatory protein that covers myosin-binding sites to prevent actin from binding to myosin
- voltage-gated sodium channels
- membrane proteins that open sodium channels in response to a sufficient voltage change, and initiate and transmit the action potential as Na+ enters through the channel
- Z-disc
- also Z-line; structure that forms the border of a sarcomere and serves as the anchor site for actin.
Contributors and Attributions
OpenStax Anatomy & Physiology (CC BY 4.0). Access for free at https://openstax.org/books/anatomy-and-physiology